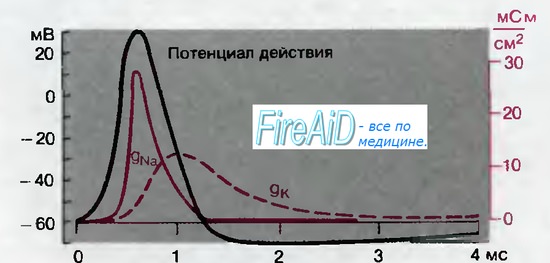
Рис. 2.7. Мембранные проводимости во время потенциала действия в гигантском аксоне кальмара. gNa и gK рассчитывали, подавая серии деполяризующих скачков потенциала (см. рис. 2.6) (по [6] с изменениями)
До сих пор мы не упоминали о том, что при деполяризации клетки открываются также Са2+-каналы. При этом возникает входящий кальциевый ток, который вместе с одновременно развивающимся Na+-током обеспечивает деполяризацию мембраны. Концентрация свободных ионов Са+ в клетке очень низка (табл. 1.1), так что равновесный потенциал для Са2 + более положителен, чем ENa. В аксонной мембране gCa меньше по сравнению с gNa, поэтому этой величиной можно пренебречь при анализе потенциала действия (рис. 2.7). Однако в дендритах нейронов или в окончаниях аксонов во время деполяризации gCa может возрастать, превышая gNa. В миокарде и тем более в гладких мышцах повышение gCa бывает столь же велико, как и повышение gNa, а иногда и более значительно. Такие входящие токи Са2+ представляют особый интерес из-за их влияния на внутриклеточную концентрацию Са2+, [Са2+];, которая может возрастать с Ю-7 до 10~6 М; это повышение [Са2 + ]; часто выполняет в клетке регулирующие функции. Механизм открывания Са2 + -каналов и последующие внутриклеточные процессы являются филогенетически очень древними-они выявлены даже у простейших.
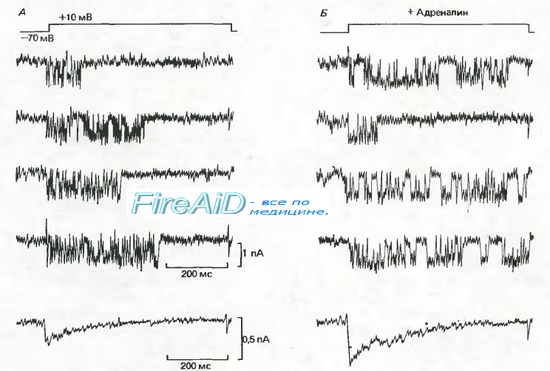
Рис. 2.14. А, Б. Токи одиночных кальциевых каналов в клетках миокарда. Вверху: деполяризация длительностью 600 мс от —70 до +10 мВ, создаваемая методом локальной фиксации потенциала. Ниже представлены 4 записи токов одиночного канала. А В нормальных условиях деполяризация в 30% случаев не вызывает токов через канал (записи не представлены). Внизу: суммарный ток, полученный путем усреднения многих индивидуальных записей токов одиночного канала; видна инактивация Са2 + -тока после деполяризации. Б. В присутствии 1 мкМ адреналина группы открываний одиночного канала становятся продолжительнее; при этом деполяризация не вызывает открываний канала только в 20%. Адреналин не влияет на амплитуду токов одиночного канала, но амплитуда суммарного тока (внизу) значительно возрастает (по [32] с изменениями)
Токи одиночных Са2 -каналов в миокарде (рис. 2.14) характеризуются несколько более сложным поведением по сравнению с Na+- и К+-токами, показанными на рис. 2.12. Во время серий деполяризационных скачков потенциала примерно в 70% случаев возникают довольно длительные вспышки импульсов тока, каждый амплитудой около 1 пА, а в 30% случаев канал остается закрытым. Индивидуальные открывания во время вспышек продолжаются в среднем около 1 мс, а закрытые состояния между ними-только 0,2 мс. Суммарный Са2+-ток во время деполяризации (нижние записи на рис. 2.14) быстро нарастает и инактивируется с постоянной времени примерно 130 мс, причем общий ток определяется длительностью и частотой вспышек. Кинетику канала проще всего описать (в соответствии с рис. 2.13) следующим уравнением:
Здесь от переходов между «Закрытым состоянием 2» и «Открытым состоянием» зависит длительность и частота индивидуальных открываний, а от переходов между «Закрытым состоянием 1» и «Закрытым состоянием 2»-частота и длительность вспышек. Уравнение (2) требует дополнения, чтобы учесть инактивированное состояние, как показано на рис. 2.13 [32].
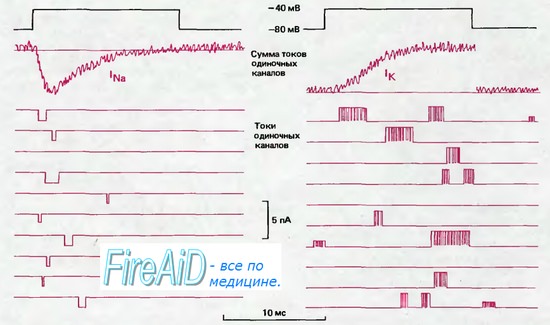
Рис. 2.12. Токи через натриевые (слева) и калиевые (справа) каналы (схематическое изображение). С помощью локальной фиксации потенциала производили сдвиг потенциала длительностью 14 мс от —80 до —40 мВ (черная линия); ниже показаны мембранные токи, зарегистрированные при нескольких таких последовательных сдвигах потенциала. Во время деполяризации токи одиночного канала могут возникать в любой момент, причем длительность их варьирует. При объединении многих записей токов в условиях синхронизации скачков потенциала получаются суммарные кривые токов, показанные вверху красным (lNa и lK). Временной ход lNa свидетельствует о том, что вероятность открывания Na+ -каналов наиболее высока вскоре после скачка потенциала, а примерно через 1 мс эти каналы открываются все реже и в конце концов инактивируются. Большая часть К+-каналов открывается с некоторой задержкой после скачка потенциала, затем средняя частота открываний остается на постоянном уровне в течение всего периода деполяризации.
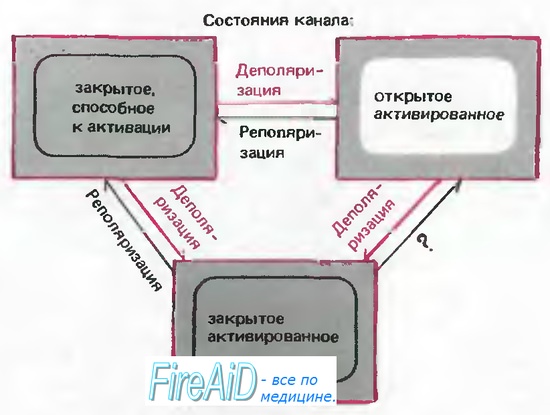
Рис. 2.13. Модель состояний Na+-каналов. «Закрытое, способное к активации» состояние при деполяризации может преобразовываться в «открытое активированное» или «закрытое инактивированное» состояние. Когда канал находится в «открытом активированном» состоянии, стойкая деполяризация способствует переходу в «закрытое инактивированное» состояние. Возвращение канала в «закрытое, способное к активации» состояние может происходить только в результате реполяризации. (Более реальная модель включает последовательно 3 «закрытых, способных к активации» и 4 «закрытых инак-тивированных» состояния [8].)
Записи активности Са2+-канала на рис. 2.14 служат также примером модуляции активности канала гормоном или медиатором. Адреналин, секретируемый корой надпочечников как «эрготропный гормон», поступает к сердцу с кровотоком; один из его эффектов состоит в увеличении частоты сердечных сокращений. Кроме того, он высвобождается (вместе с норадреналином) в качестве медиатора из симпатических нервов сердца, вызывая тот же эффект. В эксперименте, результаты которого приведены на рис. 2.14, Б, адреналин в концентрации 10 ~6 М апплицировали на клетку миокарда. После этого деполяризация вызвала примерно в 80% случаев активность одиночных Са2+-каналов с повышенной частотой вспышек. Кратковременные открывания и закрывания каналов были такими же, как раньше. Суммарная кривая (рис. 2.14, Б, внизу) отчетливо показывает, что адреналин увеличивал вход Са2+. Такой же эффект можно вызвать перфузией клеток миокарда раствором с циклическим аденозинмонофосфатом (цАМФ) или применением каталитической субъединицы цАМФ-зависимой протеинкиназы (ПК-А). Эти наблюдения свидетельствуют, что адреналин действует здесь через второй посредник-цАМФ, вызывая фосфорилирование ферментов каталитической субъединицей протеинкиназы. Таким образом, адреналин, по-видимому, увеличивает Са2+-ток путем инициации фосфорилирования Са2+-канала, которое способствует переходу из «Закрытого состояния 1» в «Закрытое состояние 2». Эффект адреналина, представленный на рис. 2.14, может служить прототипом модуляции клеточной активности гормонами или медиаторами.
В мембране, несомненно, существуют еще и Сl-каналы. Они изучены недостаточно подробно, поэтому рассматриваться здесь не будут.