Натрий (Na) и калиевая (K) проводимость во время потенциала действия.
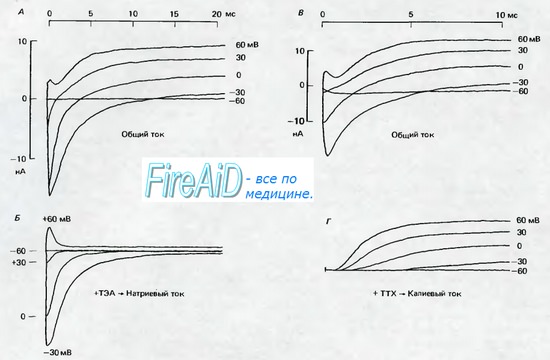
Рис. 2.6. А и В Мембранные токи в миелинизированных аксонах лягушки (перехваты Ранвье; 11 13 С) после ступенчатых сдвигов мембранного потенциала. Мембранный потенциал поддерживался с помощью фиксации потенциала на уровне потенциала покоя, равного —95 мВ; в момент времени 0 мс мембранный потенциал скачком поднимали до значений, которые указаны справа около записей тока, от —60 до +60 мВ. Сопровождающие скачок потенциала кратковременные емкостные токи вычитались, поэтому регистрируемые токи являются ионными токами. При —60 мВ скачок потенциала остается подпороговым и не вызывает изменений тока. По мере увеличения скачков потенциала сначала возникают отрицательные токи, которые с увеличением потенциала становятся положительными. Б. То же, что на рис. А, но на фоне блокады калиевых токов ТЭА (6 мМ), в результате чего токи почти полностью обеспечиваются ионами Na+. Полярность Na-токов меняется с отрицательной на положительную между значениями +30 и +60 мВ; по мере увеличения деполяризации продолжительность Na - токов уменьшается. Г. То же, что на рис. В. но на фоне блокады натриевых токов тетродотоксином (0,3 мкМ), так что записи соответствуют калиевым токам. При деполяризации К+-токи нарастают медленнее, чем Na - токи, и продолжаются в течение всего периода деполяризации (по [3] с изменениями)
Временной ход мембранной проводимости для натрия (Na) и калия(К) можно рассчитать путем деления амплитуд соответствующих токов (рис. 2.6) на разность между тестирующим и равновесным потенциалами для соответствующего иона. Подобные данные можно также получить для небольших скачков потенциала. Пусть, например, известна амплитуда тока, вызванного маленьким скачком потенциала в околопороговом диапазоне. Этот ток протекает по мембранной емкости и идет через мембранное сопротивление, значения которых известны (см. рис. 2.16 и 2.17), вызывая небольшую деполяризацию. Этот потенциал в свою очередь вызывает дополнительный ток, который ведет к дальнейшей деполяризации и значение которого можно вычислить.
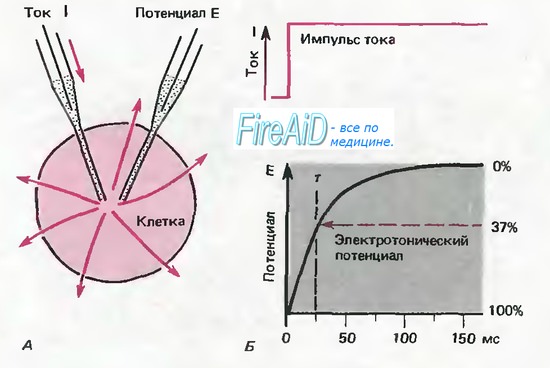
Рис. 2.16. А, Б. Электротонический потенциал в клетке сферической формы. А. Внутриклеточные электроды служат для регистрации мембранного потенциала Е и пропускания тока I, распределение которого показано красными стрелками. Б. Временной ход импульса тока и одновременно регистрируемого электротонического потенциала в клетке. Постоянная времени т электротонического потенциала определяется временем, в тече- ' ние которого потенциал доходит до уровня, достигающего 37% (1/е) его конечной амплитуды
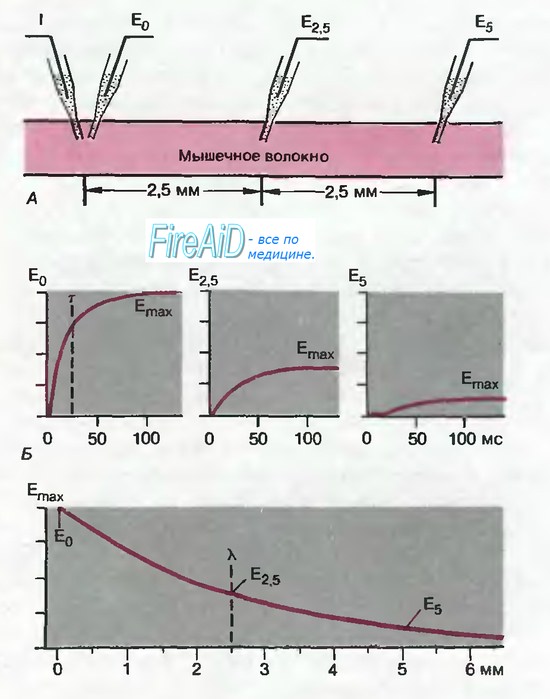
Рис. 2.17. Электротонические потенциалы в клетке вытянутой формы. Вверху: инъекция тока 2 в мышечную клетку; электротонические потенциалы регистрируются на расстояниях 0; 2,5 и Б мм. В середине: временной ход электротонических потенциалов при этих трех расстояниях; в каждом случае потенциал достигает разного конечного уровня Еmax. Внизу: зависимость Еmax( от расстояния до места инъекции тока. Постоянная длины мембраны h(лямбда) равна расстоянию, при котором Еmax падает до уровня 37% (1/е) амплитуды в месте пропускания тока
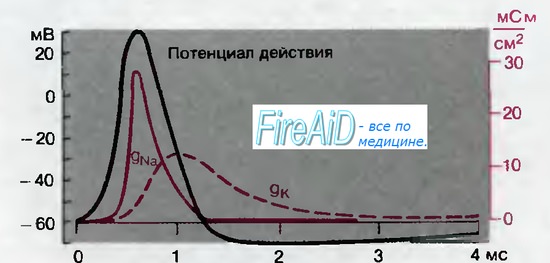
Продолжая работать с такими небольшими скачками потенциала и временными отрезками, можно воспроизвести временной ход потенциала действия по зарегистрированным потенциалзависимо-стям амплитуд и временного хода gNa и gK. На рис. 2.7 изображен реконструированный таким образом потенциал действия, а также временной ход gNa и gK. При достижении порогового потенциала gNa резко нарастает; она достигает максимума раньше пика потенциала действия, поскольку уже начинается инактивация Na+-токa, и в течение 1 мс gNa возвращается к исходному уровню. Напротив, gK нарастает медленно, с некоторой задержкой после начала деполяризации. Она достигает своего максимума поздно, когда реполяризация уже наполовину завершена, и затем снижается, так как снижается деполяризация. Таким образом, повышение gK ускоряет вторую фазу реполяризации и служит причиной гиперполяризационного следового потенциала после потенциала действия (рис. 2.7); в то время как gK все еще превышает значение потенциала покоя, мембранный потенциал смещается от уровня покоя по направлению к отрицательному калиевому равновесному потенциалу Е(k).
Рис. 2.7. Мембранные проводимости во время потенциала действия в гигантском аксоне кальмара. gNa и gK рассчитывали, подавая серии деполяризующих скачков потенциала (см. рис. 2.6) (по [6] с изменениями)