Овуляция происходит примерно через 15 ч после достижения концентрацией лютропина в плазме крови максимальных значений (пик концентрации). Лютропин связывается с рецепторами мембраны фолликулярных клеток и активирует завершение первого мейотического деления первичного ооцита (см. рис. 16.1). В результате образуется вторичный ооцит, имеющий гаплоидное число хромосом, которые во время оплодотворения яйцеклетки сливаются с гаплоидным числом хромосом сперматозоида.
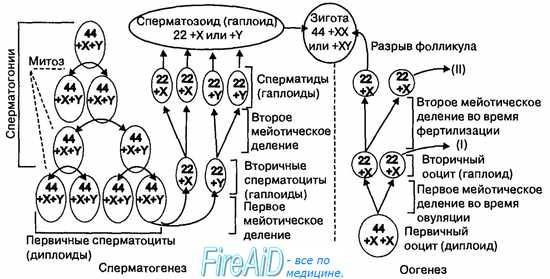
Рис. 16.1. Схема сперматогенеза и оогенеза.
Первичные сперматоциты образуются путем митоза из сперматогоний и содержат диплоидное число хромосом. В результате первого мейотического деления образуются сперматоциты, содержащие гаплоидное число двухроматидных хромосом (22 единичные соматические хромосомы и одна половая — X или Y). После второго мейотического деления образуются сперматиды, содержащие гаплоидный набор монохроматидных хромосом. Оогенез начинается со стадии первичного ооцита, содержащего диплоидное число хромосом. Первичный ооцит путем первого мейотического деления превращается во вторичный ооцит, содержащий гаплоидное число двухроматидных хромосом (22 единичные соматические хросомы и одна половая Х-хро-мосома). В процессе оплодотворения ооцита сперматозоидом образуется зигота, содержащая 22 пары соматических хромосом и одну пару половых (XX или XY). Таким образом, в зиготе восстанавливается диплоидный набор хромосом.
Кроме того, лютропин, связываясь с рецепторами системы G-белков фолликулярных клеток, увеличивает в них содержание вторичного посредника цАМФ. Под влиянием цАМФ в фолликулярных клетках непосредственно перед овуляцией прекращается фолликулогенез, связанный с образованием в гранулезных клетках стероидогенных энзимов, рецепторов фоллитропина, (3-эстрогеновых рецепторов и циклина D2, являющегося регулятором митоза в фолликулярных клетках.
Под влиянием высокой концентрации лютропина в плазме крови возрастает кровоток в капиллярах ткани фолликула за счет роста продукции NO-синтазы и образования вазодилататора оксида азота в гладких мышцах сосудов, питающих фолликул. Это увеличивает размер фолликула, стимулируя накопление фолликулярной жидкости, которая своим гидростатическим давлением истончает слой гранулезных клеток и клеток наружной оболочки (theca) настолько, что лишь эпителиальный слой, покрывающий поверхность яичника, отделяет стенку фолликула от перитонеального пространства. В предовуляторную фазу лютропин стимулирует синтез рецепторов прогестерона, пептида, активирующего синтез аденилатциклазы в гипофизе, регуляторного фактора роста, которые вызывают овуляцию.
Преовуляторное увеличение концентрации прогестерона в плазме крови женщины активирует синтез в фолликуле протеолитических энзимов и простагландинов (Е и F). Под действием простагландинов, а также тром-боксанов и лейкотриенов происходит истончение, растяжение и разрушение стенки фолликула. В результате зрелая яйцеклетка выходит через стенку лопнувшего фолликула в жидкость перитонеального простраства. Двигательная активность цилий ампулярного расширения фаллопиевой трубы направляет движение ооцита в просвет фаллопиевой трубы.