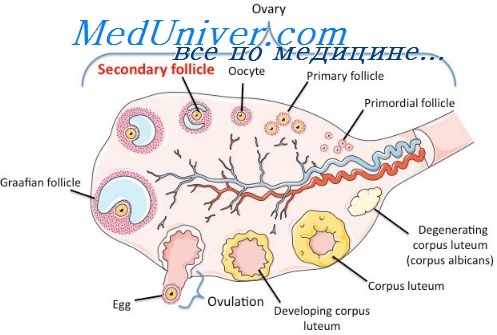
Менструальный цикл начинается с периода полового созревания (в средней полосе Европейской части России) с колебаниями от 12 до 15 лет и продолжается в детородном возрасте до 45—46 лет. Продолжительность менструального цикла в детородном возрасте у женщин с регулярными менструациями вариабельна. Примерно у 60% она составляет 28 дней и только у 10—12%—30—35 дней.
Однако современные данные свидетельствуют о том, что циклические кровянистые выделения из матки могут возникать и без овуляции. Подобные выделения называют псевдоменструацией, или менструальноподобным кровотечением.
Многочисленные экспериментальные и клинические наблюдения позволяют в настоящее время признать концепцию о иейрогуморальной регуляции менструального цикла у женщин.
Первое звено нейрогуморальной регуляции — кора головного мозга — идентично как в мужском, так и в женском организме; половые различия проявляются на уровне гипоталамуса. М. С. Мицкевич и А. Жост считают, что у человека и некоторых млекопитающих половая дифферепцировка гипоталамуса происходит уже к концу антенатального периода. Гипоталамус женского типа характеризуется тем, что сначала в нем происходит тоническая (базальная), а затем циклическая секреция гипофизотропных гормонов, гипоталамус мужского типа— только тонической секрецией гормонов без волнообразных колебаний их уровня.
Кора головного мозга. Участие коры головного мозга в регуляции функции половой системы установлено многими исследователями. Повседневные клинические наблюдения подтверждают влияние различных психогенных факторов на особенности менструальной функции. Известны случаи наступления аменореи после тяжелых психических травм.
Вместе с тем имеются некоторые данные об участии амигдалоидных (миндалевидных) ядер и лимбической системы в нейрогуморальной регуляции полового цикла. Необходимо отметить особую пластичность, способность коры головного мозга «приспосабливаться» к изменяющимся условиям внешней среды; в связи с этим обычные раздражители, как правило, не приводят к изменению половой цикличности.
Система гипоталамус — гипофиз, объединенная тесными анатомическими и физиологическими связями, представляет собой целостный функциональный комплекс и играет большую роль в регуляции менструального цикла [Алешин Б. В., Баранов В. Г. и др.].
Эти нейросекреты способны стимулировать выделение соответствующих тройных гормонов и получили ранее название разреигающих факторов (releasing factor — RF). В настоящее время установлено, что, кроме ряда факторов (гормонов), действительно стимулирующих выделение тропных гормонов, есть другие, наоборот, тормозящие их выделение. В связи с этим группу стимулирующих факторов называют либеринами, а вторую группу— статинами.
Предполагается, что местом образования этих гормонов могут служить нервные окончания (синаптосомы), в которых также обнаруживаются биогенные амины, стимулирующие выделение указанных гормонов из нейросекреторных клеток в капилляры портальной системы. Эти амины наряду с гормонами периферических эндокринных желез действуют на гипоталамус по принципу обратной связи; их рассматривают как регуляторы секреции и синтеза гипоталамических нейрогормонов.
Так, например, на содержание ПИФ в гипоталамусе влияет концентрация катехоламинов дофамина и норадрепалина, которые существенно угнетают секрецию пролактина [Сеппала М.]. Рилизинг-факторы (гормоны) можно рассматривать как своеобразные универсальные химические факторы, посредством которых осуществляется передача нервных импульсов на эндокринную систему [Юдаев Н. А.].
Регулирующая роль гипоталамуса в деятельности системы гипофиз — яичники установлена серией экспериментальных исследований с электрическим раздражением ядер гипоталамуса, имплантацией кусочков яичника и гипофиза в некоторой области гипоталамуса и использованием фармакологических агентов. По Б. В. Алешину, основным источником образования рилизинг-гормонов являются нервные клетки аркуатного и вентромедиального ядер медиобазального отдела гипоталамуса.
Многочисленные работы венгерского исследователя Flerko и соавт. показали, что фактор, реализующий секрецию лютеинизирующего гормона (люлиберин), по-видимому, вырабатывается в среднем и заднем гипоталамусе — вентромедиальном и мамиллярном ядрах.
Рилизинг-фактор фолликулостимулнрующего гормона (фоллиберин) вырабатывается в нейронах паравентрикулярных ядер переднего гипоталамуса.
Гормоны задней доли гипофиза (окситоцин, вазопрессин и антидиуретический гормон), являющиеся нейросекретом гипоталамуса, по нервным аксонам переносятся в гипофиз и накапливаются в задней его доле. В нейрогипофизе эти гормоны соединяются с белком и в связанном виде оказывают влияние на соответствующие эффекторы.
Так, Teresawa и Timiras, Teresawa и Sawyer обнаружили рецепторы ЛГ в ростральной части аркуатного ядра и nuclei medianae в районе эминесценции, а также усиление клеточной активности в этих областях при искусственно вызванной овуляции. В супраоптическом ядре обнаружены клетки, чувствительные к осмотическому давлению крови, в вентроме-диальном ядре — к содержанию глюкозы, в области медиоба-залыюго гипоталамуса — к стероидным гормонам. Рецептивными полями этой области эстрогены, прогестерон, тестостерон и кортикостероиды связываются так же интенсивно, как и рецептивными полями аденогипофиза.
Таким образом, клетки гипоталамуса выполняют функции рецепторов, воспринимающих изменения гомеостаза, и обладают способностью трансформации гуморальных изменений внутренней среды в нервный процесс.
По химической природе нейрогормоны являются комплексом аминокислот. Однако полная химическая структура установлена лишь у трех из этих гормонов — ТРФ, ЛРФ и СИФ; остальные получены лишь в частично очищенном виде. Отчасти это зависит от крайне незначительного содержания этих гормонов (нанограммы).
Механизм действия рилизинг-гормонов на клетки адеиогипофиза обусловлен деполяризацией клеточных мембран и повышением их проницаемости. Повышение Са2+ активирует движение гранул гонадотропинов к клеточным мембранам и выход их в кровяное русло [Баранов В. Г. и др.].
Для секреторных нейронов гипоталамуса типична тесная связь с сосудистым руслом и системой полостей III желудочка головного мозга.
Нейросекреты гипоталамуса оказывают биологическое действие на организм различными путями. Основным является параденогипофизарный путь — через вены, впадающие в синусы твердой мозговой оболочки, в ток крови. Меньшая часть нейрогормонов направляется трансгипофизарно, через систему воротной вены к передней доле гипофиза. Большинство авторов считают, что кровообращение в портальной вене осуществляется нисходящим путем — от центрального возвышения к дисталыюй части. Нейросекрет попадает в гипофиз и по лимфатическим путям — в ножку гипофиза. Обратное влияние на гипоталамус, так называемый стероидный контроль половых центров, осуществляется через вертебральные артерии.
Гипоталамус играет также важную роль в регуляции вегетативных функций. Ему свойственны авторегулирующие свойства, благодаря чему осуществляется интеграция вегетативных и эндокринных функций организма.
На протяжении полового цикла интенсивность обменных процессов в диэнцефальной области претерпевает циклические колебания. Наибольшее поглощение диэнцефальной областью кислорода обнаруживается соответственно периоду овуляции. Это подтверждается также экспериментами с веществами, стимулирующими овуляцию.