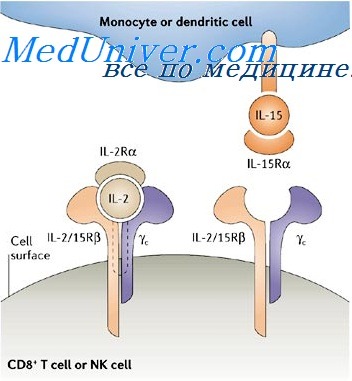
IL-15 (14-15 кДа) и IL-2 используют IL-2Rp и IL-2Ry цепи в качестве рецепторных структур.
Данные, полученные у мышей, дефицитных по генам-мишеням в индивидуальных компонентах IL-2/IL15 системы, демонстрируют различные роли IL-2 и IL-15 для активации врожденной иммунной системы. IL-15 является ключевым фактором для поддержания NK-клеточного гомеостаза, способствует дифференцировке у6Т-клеток и индукции CD8-T-клеток памяти, а также участвует в дифференцировке моноцитов в клетки Лангерганса.
Кроме того, влияние INF-a на дифференцировку и/или активацию ДК, по-видимому, опосредовано IL-15, который продуцируется ДК в ответ на IFN-a. К тому же, IL-15 является мощным ингибитором апоптоза. В присутствии IL-15 усиливаются фагоцитоз и продукция хемокинов нейтрофилами, и продукция данного цитокина значительно возрастает в ответ на множество сигналов, запускающих врожденный иммунный ответ, таких как ЛПС, микобактерии, или Toxoplasma gondii. Экзогенный IL-15 играет ключевую роль в защите хозяина от Salmonella, Plasmodium falciparum и Cryptococcus neoformans.
IL-18 (18,3 кДа) является провоспалительным цитокином, принадлежащим к семейству IL-1, благодаря сходству его структуры, рецепторного семейства и пути сигнальной трансдукции. Подобно IL-1B, IL-18 синтезируется как предшественник, требующий каспазу-1 для расщепления в активную IL-18 молекулу. Продуцируемый, главным образом, антигенпрезентирующими клетками, он индуцирует продукцию Th1 цитокинов (INF-y) и усиливает клеточно-опосредованную цитотоксичность в присутствии IL-12.
Отмечено, что IL-18 в отсутствии IL-12 индуцирует продукцию Тh2-родственных цитокинов Т- и NK-клетками, и базофилами/тучными клетками. Кроме того, IL-18 является цитокином, активирующим врожденный иммунитет и оба типа (Th1 и Th2) иммунного ответа.
IL-23 — гетеродимер, содержащий IL-12р40, и недавно клонированную IL-23 специфичную р19 субъединицу, связывающий рецепторный комплекс, состоящий из IL-12 Rp1 субъединицы и отдельного IL-23R. IL-12 и IL-23 усиливают клеточный иммунитет путем индукции синтеза INF-y и пролиферативного ответа клеток-мишеней. IL-23, продуцируемый преимущественно макрофагами и ДК, отличается от IL-12 по действию на популяцию Т-клеток. В то время как IL-12 активирует наивные и эффекторные СD4+Т-клетки, IL-23 предпочтительно действует на CD4+Т-клетки-памяти.
Недавние исследования экспрессии рецептора IL-23 и сверхэкспрессии IL-23 в трансгенных мышах выявили, что IL-23 играет критическую роль в аутоиммунном воспалении мозга, оказывая непосредственное действие на макрофагальную функцию.
IL-27 — новый открытый гетеродимерный цитокин, состоящий из IL-12р40-родственного белка, EBI-3 (Epstein-Barr virus-induced gene 3) и р28, IL-12р35-родственного полипептида. IL-27 является мощным индуктором продукции INF-y, частично в синергизме с IL-12 и IL-18. EBI-3 играет критическую регуляторную роль в индукции иммунного ответа Th2 типа, посколько у мышей, дефицитных по EBI-3 снижается количество инвариантных NKT-клеток, а также отмечается снижение IL-4 и INF-y по сравнению с нормальными мышами.
IL-27 продуцируется активированными антигенпрезентирующими клетками и в большей степени вовлекается в дифференцировку наивных CD4+Т-клеток в Тh1 тип. Напротив, IL-23 (подобно IL-15) явпяется ключевым для поддержания иммунологической памяти, потому что IL-23R экспрессируется на клетках памяти, но не на наивных Т-клетках.