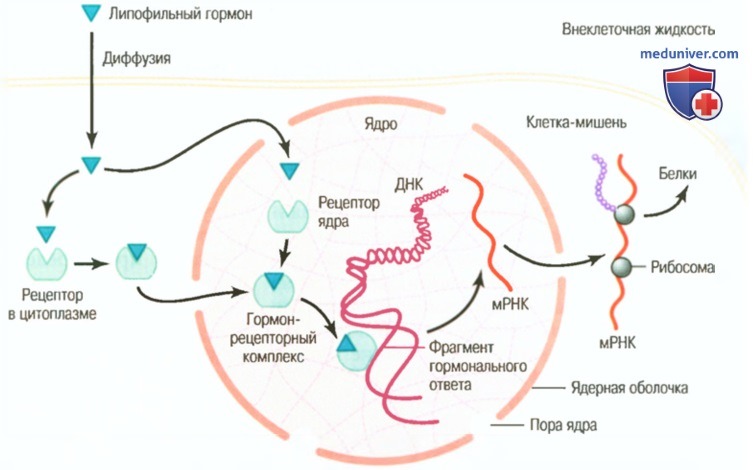
а) Внутриклеточные рецепторы гормонов и активация генов. Некоторые гормоны, в том числе стероиды коры надпочечников и гонад, гормоны щитовидной железы, ретиноидные гормоны и витамин D, связываются с белковыми рецепторами в основном внутри клетки, а не на ее поверхности. Эти гормоны жирорастворимы, поэтому легко проникают через мембрану и взаимодействуют с рецепторами в цитоплазме или ядре. Активированный гормон-рецепторный комплекс взаимодействует со специфическим регулятором (промоутером) последовательности в ДНК, называемым элементом гормонального ответа.
Таким образом, он активирует или репрессирует транскрипцию конкретных генов и образование матричной РНК (для облегчения понимания просим вас изучить рисунок ниже), поэтому через несколько минут, часов и даже дней после поступления гормона в клетку в ней появляются вновь сформированные белки и становятся регуляторами новых или измененных функций клетки.
Механизмы взаимодействия липотропных гормонов, таких как стероиды, с внутриклеточным рецептором клеток-мишеней. После взаимодействия гормона с рецептором в цитоплазме или ядре гормон-рецепторный комплекс присоединяется к фрагменту гормонального ответа (пособнику) на ДНК, что активирует или тормозит транскрипцию гена, образование матричной РНК и синтез белка
Многие ткани имеют идентичные внутриклеточные гормональные рецепторы, однако гены, регулируемые этими рецепторами, различны. Внутриклеточные рецепторы могут активировать генный ответ только в случае наличия в клетке соответствующих комбинаций ген-регуляторных белков. Многие из этих протеин-регуляторных комплексов имеют в разных тканях свои особенности, поэтому ответ различных тканей предопределяется не только специфичностью рецепторов, но также генами, которые регулируются через эти рецепторы.
б) Механизмы вторичных посредников, осуществляющих передачу гормональных влияний внутри клетки. Ранее мы отметили один из способов, с помощью которого гормоны вызывают ответы клеток и стимуляцию образования вторичного посредника цАМФ внутри клетки. Затем цАМФ становится причиной запуска последовательных внутриклеточных ответов на действие гормона. Итак, непосредственное действие гормона на клетку заключается в активации индуцирующего рецептора на мембране, а вторичные посредники обеспечивают остальные реакции.
цАМФ — не единственный вторичный посредник, используемый гормонами. Существуют два других, наиболее важных посредника:
(1) ионы кальция, сопряженные с кальмодулином;
(2) фосфолипидные фрагменты мембран.
в) Система аденилатциклаза-цАМФ как вторичный посредник. В таблице ниже указаны некоторые из многочисленных гормонов, использующих систему аденилат-циклаза-цАМФ как механизм для стимуляции тканей-мишеней, а на рисунке ниже продемонстрирована система аденилатциклаза-цАМФ как вторичный посредник.
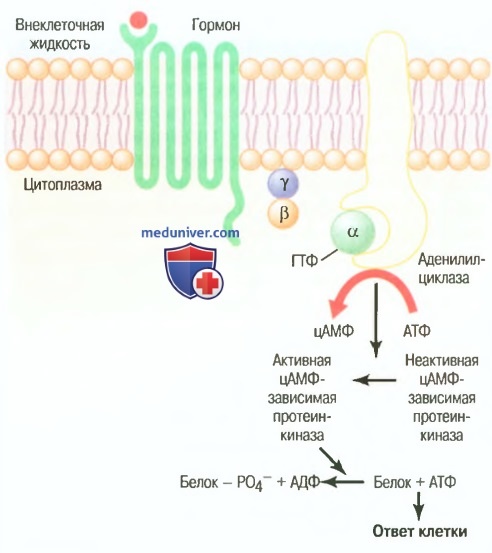
Механизм, использующий циклический аденозинмонофосфат, как способ гормональной регуляции функций клетки
Присоединение гормона к рецептору позволяет последнему взаимодействовать с G-белком. Если G-белок активирует систему аденилатциклаза-цАМФ, его называют Gs-белком, указывая на стимулирующую роль G-белка. Стимуляция аденилатциклазы, связанной с мембраной фермента посредствам Gs-белка, катализирует превращение небольшого количества присутствующего в цитоплазме аденозинтрифосфата в цАМФ внутри клетки.
Следующий этап опосредован активацией цАМФ-зависимой протеинкиназой, которая фосфорилирует специфические белки в клетке, запуская биохимические реакции, что гарантированно обеспечивает ответ клетки на действие гормона.
Как только цАМФ образуется в клетке, это обеспечивает последовательную активацию ряда ферментов, т.е. каскадную реакцию. Таким образом, первый активированный фермент активирует второй, который активирует третий. Задача такого механизма заключается в том, что небольшое количество молекул, активированных аденилатциклазой, может активировать значительно большее количество молекул на следующем этапе каскадной реакции, что является способом усиления ответа.
В итоге благодаря этому механизму ничтожно малое количество гормона, действующее на поверхность мембраны клетки, запускает мощный каскад активирующих реакций.
Если гормон взаимодействует с рецептором, сопряженным с тормозящим G-белком (Gi-белок), это снижает образование цАМФ и, как следствие, снижает активность клетки. Следовательно, в зависимости от взаимодействия гормона с рецептором, сопряженным с активирующим или тормозящим G-белком, гормон может как увеличивать, так и уменьшать концентрацию цАМФ и фосфорилирование ключевых белков клетки.
Специфичность эффекта, наблюдаемого в ответ на увеличение или уменьшение цАМФ в различных клетках, зависит от природы внутриклеточных механизмов: некоторые клетки имеют один набор ферментов, другие — иной. В связи с этим реакции, вызываемые в клетках-мишенях, разнообразны. Например, инициация синтеза специфических химических соединений вызывает сокращение или расслабление мышц либо процессы секреции в клетках или изменение проницаемости мембран.
Клетки щитовидной железы, активированные цАМФ, образуют метаболические гормоны — тироксин или трииодтиронин, в то время как тот же цАМФ в клетках надпочечников приводит к синтезу стероидных гормонов коры надпочечников. В клетках тубулярного аппарата почек цАМФ повышает проницаемость для воды.