Координация сократительной деятельности. Медленные ритмические колебания. Продольный мышечный слой. Влияние катехоламинов на миоциты.
Координация сократительной деятельности различных участков мускулатуры пищеварительной трубки осуществляется за счет регулирующих влияний периферической и центральной нервной системы. Парасимпатические нервные волокна в основном усиливают моторику желудка и кишечника. Однако в составе блуждающих нервов имеются и волокна, тормозящие моторику миоцитов. Симпатические нервы оказывают преимущественно тормозные влияния на мышечные пучки.
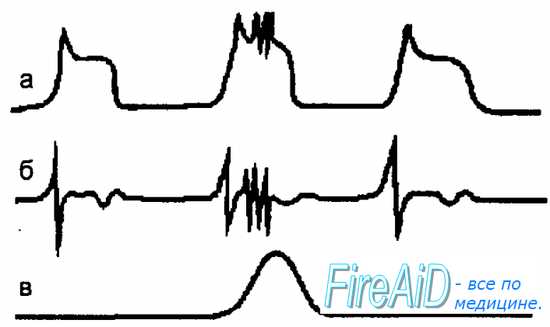
Миоциты обладают способностью генерировать медленные ритмические колебания мембранного потенциала. В фазе деполяризации мембраны происходит накопление ионов Са2+ в клетке, что активирует кальций-зависимые калиевые каналы. Это вызывает выход К+ из клетки и приводит к деполяризации мембраны. Если деполяризация достигает критического уровня, то на гребне медленной волны возникают потенциалы действия, которые вызывают открытие быстрых потенциалозависимых кальциевых каналов, что приводит к сокращению миоцита (рис. 11.3). На этом рисунке приведена синхронная запись медленных и быстрых колебаний мембранного потенциала миоцита тонкой кишки при внутриклеточной и внеклеточной регистрации. С появлением серии быстрых потенциалов возникает сокращение мышечного волокна.
Рис. 11.3. Связь между медленными и быстрыми электрическими волнами миоцита и сокращением мышечного волокна тонкой кишки. а — внутриклеточная регистрация медленных и быстрых потенциалов миоцита (потенциалы действия накладываются на плато волны деполяризации); б — внеклеточная регистрация потенциалов миоцита (серия спайковых потенциалов совпадает с медленной волной, регистрируемой при внутриклеточном отведении); в — локальное сокращение гладкого мышечного волокна.
Медленные волны деполяризации возникают в миоцитах наружной части циркулярного слоя и распространяются как на круговой, так и на продольный мышечные слои (по мышечным мостикам). Это определяет последовательность сокращения мышечных пучков, расположенных в разных мышечных слоях. Мембрана миоцитов в области расположения водителей ритма обладает способностью генерировать медленные волны с более высокой частотой, чем в соседних участках. Это способствует «навязыванию» частоты медленных волн клеток водителя ритма миоцитам соседних участков.
Продольный мышечный слой имеет выраженную холинергическую иннервацию, а циркулярный слой испытывает тонические влияния интрамуральных тормозных нейронов через их многочисленные пептидергические окончания. Нейромедиатор окончаний парасимпатических нервных волокон (ацетилхолин), действуя на М-хо-линорецепторы, вызывает повышение проницаемости клеточной мембраны к ионам Са2+, Na+ и К+. При этом увеличивается количество потенциалов действия, длительность и амплитуда фазного сокращения гладкой мышцы, что лежит в основе усиления моторики желудка и кишечника.
При сильном возбуждении парасимпатических постганглионарных нейронов повышается концентрация ацетилхолина во внеклеточной среде. Это приводит к непрерывной генерации миоцитами потенциалов действия, которые сопровождаются слиянием фазных сократительных эффектов в слитное сокращение.
Катехоламины влияют на миоциты через адренорецепторы двух типов (альфа и бета), которые подразделяются на четыре подтипа (а, и а2, B1 и B2). Воздействие норадреналина на а- и (B-адренорецепторы окончаний хо-линергических нейронов миэнтерального и подслизистого сплетений угнетает выход ацетилхолина из холинергических окончаний, что ослабляет парасимпатические влияния на миоциты и способствует торможению гладкой мускулатуры.
При электрическом раздражении симпатических нервных волокон имеет место торможение сократительной активности кишечника и усиление тонуса циркулярных пучков сфинктеров.