Торможение и возбуждение ядер мозжечка. Физиология сигнальной системы мозжечка
а) Баланс между возбуждением и торможением на уровне глубоких ядер мозжечка. Обратившись снова к контуру на рисунка ниже, следует отметить, что прямая стимуляция клеток глубоких ядер лазающими и мшистыми волокнами возбуждает их.
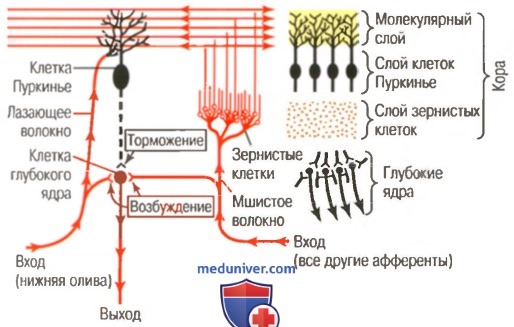
В левой стороне рисунка показан главный нервный контур мозжечка, в котором возбуждающие нейроны обозначены красным цветом, а клетка Пуркинье (тормозной нейрон) - черным цветом. Справа показана анатомическая связь глубоких ядер мозжечка и коры мозжечка с ее тремя слоями
И наоборот, сигналы, приходящие от клеток Пуркинье, тормозят их. В норме баланс между этими двумя эффектами несколько смещен в сторону возбуждения, и в условиях покоя активность клетки глубокого ядра остается относительно постоянной — на уровне умеренного непрерывного возбуждения.
При выполнении быстрого движения инициирующий его сигнал из моторной коры большого мозга или мозгового ствола сначала значительно усиливает возбуждение клеток глубоких ядер. Затем через несколько миллисекунд прибывают тормозные сигналы обратной связи от контура клеток Пуркинье. Таким образом, существует начальный быстрый возбуждающий сигнал, посылаемый клетками глубоких ядер к двигательному нисходящему пути для усиления движения, а вслед за этим в пределах небольшой доли секунды появляется тормозной сигнал.
Этот тормозной сигнал похож на сигнал отрицательной обратной связи «линии задержки», который эффективен в обеспечении демпфирования. Это значит, что при возбуждении двигательной системы после короткого периода задержки возникает сигнал отрицательной обратной связи, предупреждающий слишком сильное движение (иначе произойдет колебание движения).
б) Другие тормозные клетки в мозжечке. Кроме клеток глубоких ядер, зернистых клеток и клеток Пуркинье в мозжечке есть еще два типа нейронов: корзинчатые клетки и звездчатые клетки. Это тормозные клетки с короткими аксонами. И корзинчатые, и звездчатые клетки локализуются в молекулярном слое коры мозжечка, располагаясь среди тонких параллельных волокон, которые их стимулируют.
В свою очередь, эти клетки посылают аксоны под прямым углом сквозь параллельные волокна к прилежащим клеткам Пуркинье, вызывая их латеральное торможение, что увеличивает четкость сигнала так же, как латеральное торможение усиливает контраст сигналов во многих других контурах нервной системы.
в) Выходящие из мозжечка сигналы действуют по принципу «включение-выключение» и «выключение-включение». Типичной функцией мозжечка является участие его в обеспечении быстрых сигналов «включения» для мышц-агонистов и одновременно реципрокных сигналов «выключения» для мышц-антагонистов в начале движения. Затем при завершении движения мозжечок в основном отвечает за подачу своевременных и эффективных сигналов «выключения» к мышцам-агонистам и сигналов «включения» к мышцам-антагонистам. Все детали этого процесса пока не известны, но на основании главного мозжечкового контура (для облегчения понимания просим вас изучить рисунок выше) можно представить, как функционирует этот механизм.
Предположим, что механизм «включение-выключение» сокращения мышц-агонистов и мышц-антагонистов в начале движения начинается с сигналов из коры большого мозга. Эти сигналы направляются через немозжечковые пути мозгового ствола и спинного мозга непосредственно к мышце-агонисту, запуская начало сокращения.
В то же время параллельные сигналы поступают через мшистые волокна моста в мозжечок. Одна ветвь каждого мшистого волокна идет непосредственно к клеткам зубчатого или других глубоких ядер мозжечка. Нейроны этих ядер мгновенно посылают сигнал возбуждения назад в кортикоспинальную двигательную систему либо через таламус к коре большого мозга, либо через нервный контур мозгового ствола, чтобы поддержать сигнал, стимулирующий мышечное сокращение, которое уже было запущено корой большого мозга.
В результате через несколько миллисекунд сигнал включения становится сильнее, чем на старте, из-за суммации корковых и мозжечковых сигналов. Так происходит при нормально функционирующем мозжечке, но при отсутствии его вторичный дополнительный поддерживающий сигнал исчезает. Эта поддержка мозжечка делает включенное мышечное сокращение гораздо более сильным, чем оно было бы без мозжечка.
Теперь о том, что вызывает сигнал выключения мышц-агонистов для завершения движения. Вспомните, что все мшистые волокна имеют вторую ветвь, которая проводит сигналы через гранулярные клетки к коре мозжечка и в итоге — через параллельные волокна к клеткам Пуркинье. Клетки Пуркинье, в свою очередь, тормозят клетки глубоких ядер. Этот путь используют параллельные волокна молекулярного слоя коры мозжечка — одни из самых тонких медленнопроводящих волокон в нервной системе, диаметр которых составляет лишь долю микрометра.
Кроме того, сигналы этих волокон слабые, поэтому требуется определенный период времени для создания достаточного возбуждения в дендритах клетки Пуркинье, чтобы возбудить ее. Но как только клетка Пуркинье возбуждается, она, в свою очередь, посылает сильный тормозной сигнал к той же клетке глубокого ядра, которая сначала включила движение. Следовательно, это помогает выключить движение через короткое время.
Становится понятно, как полный контур мозжечка может вызвать быстрое включение сокращения мышцы-агониста в начале движения и, кроме того, вызвать в точно выверенный момент выключение сокращения той же мышцы-агониста.
Теперь рассмотрим функцию контура для мышц-антагонистов. Вспомним, что в спинном мозге на всем его протяжении существуют контуры реципрокных связей мышц-агонистов и мышц-антагонистов буквально для каждого движения, возникающего на уровне спинного мозга. Следовательно, эти контуры являются базой для выключения мышц-антагонистов в начале движения и включения их при завершении движения, что зеркально отражает события, происходящие с мышцами-агонистами. Но следует вспомнить также, что мозжечок содержит несколько типов тормозных клеток, кроме клеток Пуркинье.
Функции некоторых из них еще окончательно не определены; они, вероятно, играют роль в торможении мышц-антагонистов в начале движения и последующего возбуждения в конце движения.
Все эти механизмы пока отчасти спекулятивны. Они представлены здесь в основном для иллюстрации путей, с помощью которых мозжечок мог бы вызывать усиление сигналов включения и выключения, управляющих мышцами-агонистами и мышцами-антагонистами, и контролировать также развитие движения во времени.
Видео физиология торможения в ЦНС - профессор, д.м.н. П.Е. Умрюхин