Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
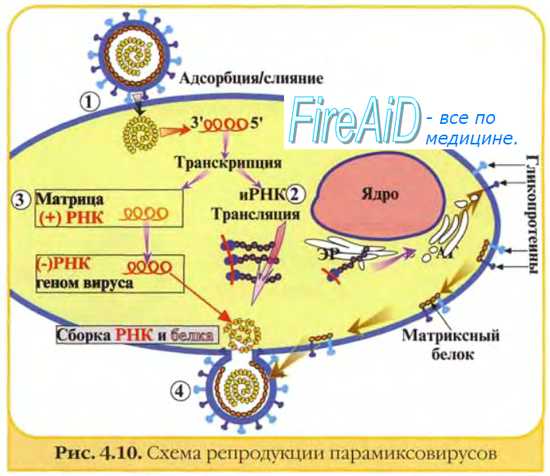
-РНК-вирусы проникают в клетку путём слияния (парамиксовирусы) либо виропексиса (рабдо- и ортомиксовирусы). Для эффективной репродукции вирусная -РНК должна быть преобразована в +РНК — аналог клеточной мРНК (рис. 5-3).
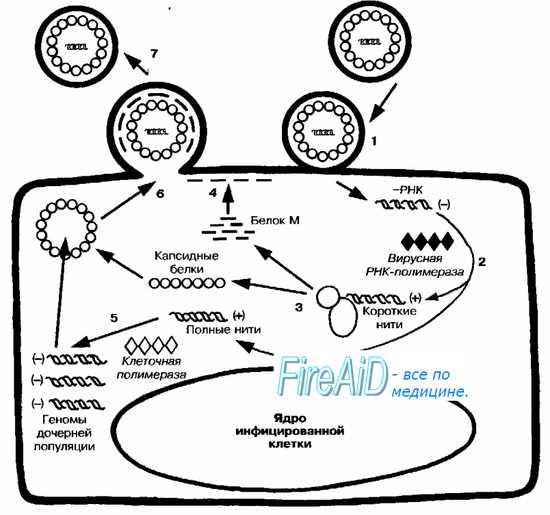
Рис. 5-3. Репродуктивный цикл -РНК-содержащих вирусов. Проникновение вируса в клетку происходит после его адсорбции и слияния с клеточной оболочкой (1). После высвобождения вирусной -РНК происходит синтез +РНК на матрице -РНК, катализируемый РНК-зависимой РНК-пол имеразой, входящей в состав вириона (2), что приводит к образованию полных и коротких нитей. Короткие +РНК-нити участвуют в синтезе ферментов и белков для дочерних популяций (3). Среди последних особую значимость имеют белок М (4) и гликопротеины оболочки, встраивающиеся в клеточную стенку на этапах, предшествующих отпочковыванию. Полная цепь +РНК служит матрицей для синтеза молекул -РНК дочерних популяций (5). Вирионы дочерних популяций собираются на участках клеточной мембраны, модифицированных белком М (6), и высвобождаются почкованием, захватывая её фрагмент, служащий в дальнейшем суперкапсидом (7).
-РНК-вирусы. Репродукция -РНК-вирусов
Ранняя стадия репродукции. После высвобождения генома вирусная транскриптаза (РНК зависимая РНК-полимераза) запускает синтез +РНК. При этом «шаблоном» для вирусной транскриптазы служит вирусный рибонуклеопротеин (то есть РНК и внутренние белки) В результате образуются полные и короткие молекулы-копии +РНК.
Поздняя стадия репродукции. Полные плюс-нити служат матрицами для синтеза молекул -РНК, составляющих геномы дочерней популяции. Короткие плюс-нити участвуют в синтезе ферментов и белков. Вирусные белки (гемагглютинин и нейраминидаза) взаимодействуют участками клеточной мембраны. Там же сорбируются и вирусные М-белки (белки матрикса) Они проявляют выраженную гидрофобность за счёт содержания до 75% нейтральных аминокислот. Это свойство даёт им способность взаимодействовать с белками и липидами клеточные мембраны и быть посредником сборки вирусных частиц. С одной стороны, М-белок распознает участки включения гликопротеинов вируса в мембрану, с другой — его специфически распознает нуклеокапсид и связывается с ним. Сборка дочерних популяций завершается после присоединения нуклеокапсида к клеточной мембране. Их высвобождение происходит путём почкования через модифицированные участки мембраны. Отпочковывающиеся вирусные частицы захватывают её фрагменты, служащие в дальнейшем суперкапсидами.
Репродукция вирусов с двухнитевыми РНК
Двухнитевые РНК-вирусы представлены семейством Reoviridae (рео- и ротавирусы). Они не имеют суперкапсида и организованы по типу кубической симметрии. С вирусной РНК связана РНК-зависимая РНК-полимераза. Вирусы отличает удлинённый репродуктивный цикл и тенденция к накоплению продуктов вирусспецифического синтеза внутри клеток. После высвобождения генома в цитоплазме клеток РНК-полимераза осуществляет синтез молекул мРНК (+РНК на одной нити -РНК. В результате образуется до 11 функциональных молекул мРНК, соответствуюших по размерам 11 сегментам одной нити -РНК. Молекулы транслируются в 11 первичных полипептидных продуктов. Их последующее расщепление приводит к образованию в заражённых клетках до 16 вторичных полипептидов. Семь первичных и два вторичных полипептида входят в состав вирусных частиц, остальные первичные и вторичные полипептиды выполняют каталитические и регуляторные функции. Параллельно, синтезированная в ходе трансляции вирусная РНК-полимераза запускает синтез минус-нитей на матрице +РНК с последующим их соединением в двухнитевую молекулу РНК. Выход образовавшихся вирионов сопровождается гибелью клетки.