Катаболизм азотсодержащих органических соединений бактериями. Аминокислоты. Декарбоксилирование и дезаминирование аминокислот бактериями. Механизм Стиклэнда.
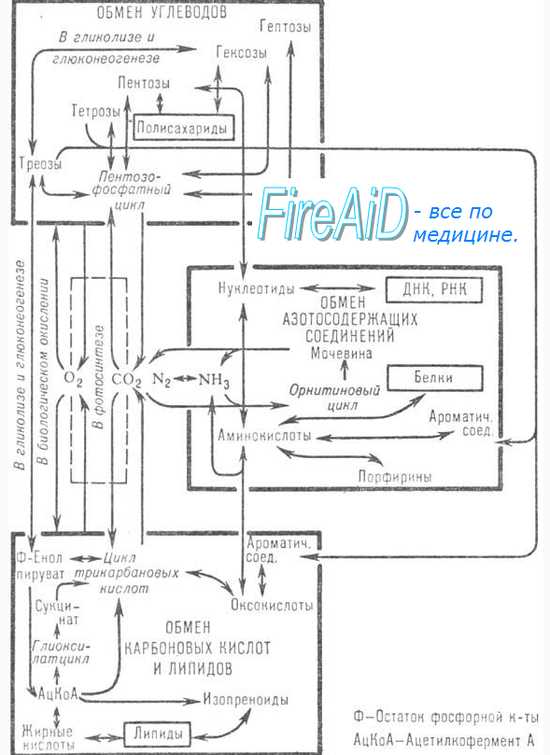
Доступные субстраты для получения углерода, азота и энергии — аминокислоты, пурины и пиримидины. Как аэробные, так и анаэробные бактерии используют эти соединения для синтеза белка либо непосредственно, либо после ряда превращений и вовлечения в промежуточный обмен.
Аминокислоты. Декарбоксилирование и дезаминирование аминокислот бактериями. Механизм Стиклэнда.
Декарбоксилирование и дезаминирование аминокислот. Первой реакцией катаболизма аминокислот может быть декарбоксилирование либо дезаминирование. Декарбоксилазы действуют обычно в кислой среде, образуя С02 и первичные амины (так называемые биогенные амины, трупные яды — кадаверин, путресцин, агматин). Поскольку при этом высвобождаются основные группы (амины), то такой процесс рассматривают как механизм нейтрализации среды и сохранения рН в физиологических пределах.
Дезаминирование аминокислот идёт с выделением аммиака. В зависимости от судьбы углеродного скелета различают дезаминирование окислительное (наиболее распространённое, например, превращение глутаминовой кислоты в 2-оксоглутаровую), гидролитическое и приводящее к образованию ненасыщенных соединений. Ферменты, катализирующие эти реакции, обычно специфичны для D- и L-изомеров аминокислот. Углеродные фрагменты, не содержащие азота, используются в процессах брожения или дыхания. Если в состав аминокислот входит сера, то последняя обычно высвобождается в форме сероводорода или меркаптанов. Разложение ароматических аминокислот (например, триптофана) происходит с образованием индола и скатола. У некоторых микроорганизмов в качестве источников энергии могут использоваться лишь некоторые продукты дезаминирования. Например, эшерихии и протеи дезаминируют триптофан с образованием индола и пирувата, из которых лишь последний утилизируется как источник энергии. Поскольку индол накапливается в культуре, то его наличие легко обнаруживают с помощью реактива Эрлиха (смесь л-диметиламинобензальдегида и НСl в этаноле), что используют для идентификации бактерий на практике.
• Некоторые бактерии обладают специальными механизмами получения энергии при расщеплении аминокислот. Например, аргинин расщепляет аргининдегидролазная система, состоящая из нескольких ферментов. Первоначально аргининдезаминаза катализирует его превращение в цитрул-лин, затем последний превращается в орнитин через реакцию, сопряжённую с синтезом АТФ.
• На средах, содержащих смесь аминокислот, многие клостридии получают большую часть энергии не из отдельных компонентов, а путём сопряжения окислительно-восстановительных реакций между парами подходящих аминокислот, известного как механизм Стиклэнда. С этих позиций аминокислоты можно разделить на акцепторы (глицин, орнитин, пролин) и доноры водорода (аланин, изолейцин и валин). Первоначально донор окисляется до кетокислоты, затем «доокисляется» до жирной кислоты. Образующийся при этом НАДН+ утилизируется для восстановления другой аминокислоты — акцептора (или, реже, другого азотистого соединения).
Переаминирование аминокислот. Кроме реакций дезаминирования и декарбоксилирования, аминокислоты могут подвергаться переаминированию, то есть переносу целой аминогруппы от аминокислоты к а-кетокислотам без промежуточного образования аммиака. Участвующая в переамини-ровании аминокислота (донор аминогруппы) превращается в а-кетокислоту {продукт окислительного дезаминирования), а сс-кетокислота (акцептор) подвергается восстановительному аминированию. Реакции катализируют специфические трансферазы. В реакциях переаминиро-вания участвуют все L-аминокислоты, при этом на а-кетокислоты переносятся аминогруппы не только в а-положении, но и в других положениях.
Пурины и пиримидины
Пурины и пиримидины становятся доступными для энергетического метаболизма лишь после гидролиза нуклеотидов и нуклеозидов. В результате их разложения образуются углекислота, аммиак, муравьиная, уксусная и молочная кислоты, часть из которых включается в рассмотренные выше энергетические пути.