Некоторые палочковидные бактерии могут образовывать нитевидные формы, особенно в неблагоприятных условиях. Формирование подобных структур осуществляется путём соединения отдельных клеток с помощью слизи, мостиков или специальных полисахаридных футляров. Многие патогенные бактерии способны также образовывать нитевидные и даже ветвящиеся структуры при нарушениях условий роста и регуляции деления. После нормализации условий существования бактерии восстанавливают нормальные форму и размеры (реверсируют к исходному фенотипу). Следует помнить, что подобные инволюционные формы особенно часто наблюдают при культивировании на искусственных питательных средах либо при проведении антибактериальной терапии.
Бактерии размножаются поперечным (бинарным) делением с образованием двух идентичных особей. Делению предшествует удвоение бактериального генома, известное как репликация. Каждая цепь двойной спирали ДНК представляет собой линейную полярную последовательность дезоксирибонуклеотидов, расположенных в строго определённом порядке. Полярность цепи связана с тем, что каждый нуклеотид имеет 5'-фосфатный конец и З'-гидроксильный конец. При соединении нуклеотидов образуется цепь, также имеющая 5'-фосфатный и 3'-гидроксильный конец, причём цепи двойной спирали имеют противоположную полярность, то есть антипараллельны (рис. 2-6). Информационное содержание обеих цепей идентично (цепи гомологичны, или комплементарны), так как каждая из них содержит последовательность нуклеотидов, строго соответствующую последовательности другой цепи благодаря образованию водородных связей между парами аденин-тимин и гуанин-цитозин.
Репликация начинается в определённой точке ДНК и осуществляется ферментами ДНК-полимеразам и одновременно на каждой матричной цепи в двух противоположных направлениях, поскольку ДНК-полимеразы могут считывать матрицу только в направлении 3'-5'. При этом на одной из матричных цепей синтез ДНК происходит непрерывно (образуется ведущая дочерняя цепь), а на другой — короткими фрагментами длиной около 100 нуклеотидов, названными по имени открывшего их учёного фрагментами Оказаки (отстающая цепь). Синтез всегда начинается с РНК-затравки, которая затем удаляется, а фрагменты сшиваются лигазами (рис. 2-7). В результате репликации число молекул ДНК в клетке удваивается. Вновь синтезированные молекулы ДНК постепенно расходятся в образующиеся дочерние клетки. Всё это позволяет дочерней клетке иметь тождественную материнской клетке молекулу ДНК. Считают, что репликация ДНК занимает почти 80% времени, в течение которого осуществляется деление бактериальной клетки.
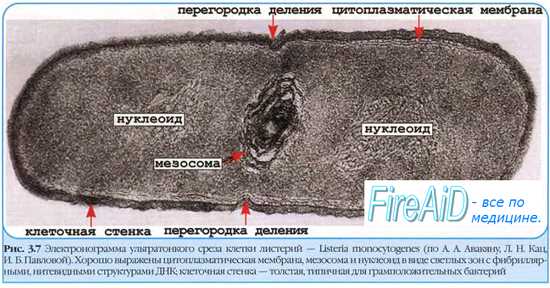
После завершения репликации ДНК начинается образование межклеточной перегородки; вначале с обеих сторон клетки происходит врастание двух слоев ЦПМ, а затем между ними синтезируется пептидогликан и образуется перегородка, состоящая из двух слоев ЦПМ и пептидогликана. Во время репликации ДНК и образования перегородки бактериальная клетка непрерывно растёт. В этот период происходят синтез пептидогликана клеточной стенки, ЦПМ, образование новых рибосом и других органелл и соединений, входящих в состав цитоплазмы.
На последней стадии деления дочерние клетки отделяются друг от друга. Процесс разделения клеток у некоторых бактерий идёт не до конца; в результате образуются цепочки клеток. При делении грамположительных бактерий образуется перегородка, отрастающая от клеточной стенки к центру клетки. У видов рода Mycobacterium перегородка образуется внутри клетки, затем расщепляется на два слоя и разделяет одну клетку на две. Грамотрицательные бактерии (включая риккетсии) истончаются в центре и разделяются перегородкой на две клетки. Бактерии рода Francisetla размножаются способом, напоминающим почкование у дрожжей (см. далее).