Ретровирусы. Семейство ретровирусов. Виды ретровирусов.
Ретровирусы содержат две идентичные молекулы геномной РНК и РНК-зависимую ДНК-полимеразу (обратную транскриптазу, ревертазу). Ретровирусы выделены от множества видов животных и проявляют разнообразный спектр патогенного потенциала. Семейство включает вирусы 7 родов: альфа-, бета-, гамма-, дельта-, эпсилонретровирусы, лентивирусы и спумавирусы. Семейство включает вирусы, патогенные для человека и многих видов животных.
Большинство ретровирусов обладают выраженным тропизмом к клеткам лимфОретикулярной и гемопоэтической системы. Ретровирусы у специфических для них видов животных встречаются повсеместно. В борьбе с ретровирусными инфекциями основной мерой является предотвращение передачи вируса.
Вирионы ретровирусов представляют собой округлые оболочечные частицы диаметром 80—100 нм, обладающие уникальной трехслойной структурой. Центральная часть вириона представлена нуклеопротеиновым комплексом, который включает около 30 молекул ревертазы и имеет спиральную симметрию. Эта структура окружена икосаэдрическим капсидом диаметром около 60 нм, покрытым оболочкой, происходящей из мембраны клетки, от которой отходят гликопротеиновые пепломеры. Лентивирусы имеют на поверхности примерно 72 шишкоподобных пепломера длиной около 10 нм с яйцевидным концевым уплотнением.
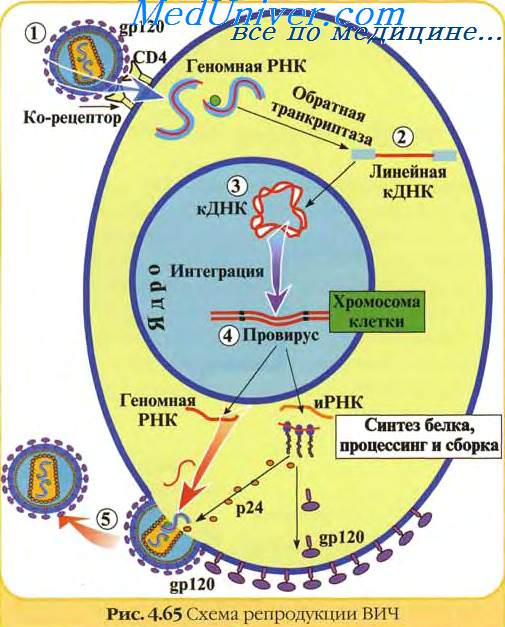
Ретровирусы имеют диплоидный геном, представляющий инвертированный димер из двух молекул линейной позитивной полярности, оцРНК; каждая молекула содержит 7-11 тн и имеет полиА последовательность на З'-конце и КЭП-структуру на 5'-конце. Детальная организация геномов разных ретровирусов широко варьирует.
Геном ретровирусов уникален в следующих отношениях:
1) является единственным диплоидным;
2) вирусная РНК синтезируется и изменяется с помощью механизма, изменяющего клеточную мРНК;
3) это единственный геном, связанный со специфическим переносом функции РНК целиком к первичной репликации;
4) это единственная оц(+)РНК, которая не является мРНК вскоре после инфекции;
5) это единственный геном, кодирующий обратную транскриптазу, которая сама по себе уникальна.
Среди многих своих функций обратная транскриптаза служит в качестве РНК-зависимой ДНК полимеразы, а ДНК-зависимая ДНК полимераза, интеграза и РНКаза, каждая в меру своей определенной функции, представляют различную часть белковой молекулы. Геном недефектных ретровирусов содержит три разных гена, каждый из которых кодирует два и более белка. Gag ген кодирует вирионные коровью белки, pol ген кодирует обратную транскриптазу, a env ген кодирует вирионные пепломерные белки. Геном кодирует также несколько определенных вирионных компонентов.
Геном быстро трансформирующих ретровирусов содержит четвертый ген — вирусный онкоген (v-onc). Присутствие онкогена обычно связано с делецией в другом месте генома, обычно в env гене, так что большинство v-onc содержащих вирусов не способны синтезировать полностью оболочку и поэтому являются дефектными по репликации. Они всегда находят связь с недефектными вирусами, которые реплицируются полностью и функционируют в качестве помощников. Вирус саркомы Рауса является исключением. Его геном содержит вирусный онкоген v-src и также полные gag, pol и env гены, и поэтому является репликативнокомпетентным.
Кроме генов gag, pol и env, общих для всех ретровирусов, лентивирусы кодируют еще несколько других «дополнительных» генов. К ним относятся:
1) tat ген, который кодирует трансактиватор, связанный с клеточными факторами, повышающими эффективность трансляции клеточной РНК полимеразы, главным образом за счет преждевременной остановки транскрипции;
2) rev ген кодирует белок, который связан с удлинением РНК транскриптов и их перемещением из ядра в цитоплазму, обеспечивая полный путь мРНК для трансляции;
3) nef ген, который не требуется для репликации вируса в культуре лимфоцитов, но важен для репликации в макрофагах и для развития болезни, вызванной вирусом иммунодефицита обезьян у обезьян резус; nef белок регулирует экспрессию клеток СД4 и образование ИЛ-2 и может изменять активацию клеток мишеней в организме;
4) vif ген кодирует белок (фактор инфекционности вирусов), который необходим для некоторых стадий вирусного морфогенеза, связанных с инфекционностью.
Vpu, vpr и vpx найдены в лентивирусах приматов. Vpu способствует созреванию вирусных гликопротеинов и освобождению вирионов почкованием; Vpr -слабый активатор транскрипции; Vpx, по-видимому, способствует репликации в Т-лимфоцитах и макрофагах.
При продуктивной инфекции вирионы формируются и освобождаются почкованием через плазматическую мембрану. Некоторые ретровирусы вызывают образование опухолей.
Спумавирусы являются необычными ретровирусами во многих отношениях. Они выделены от человека, обезьян, КРС, кошек, морских львов и хомяков. Они представляют проблему, когда контаминируют клеточные культуры, но их связь с заболеваниями неизвестна. Существует мнение, что пенящие вирусы ближе стоят к ДНК гепаднавирусам.
Эндогенный ретровирус свиней (PERV) найден у всех исследованных пород свиней. Известны три субгруппы этого вируса: А, В и С. Все свиньи содержат множественные провирусы (около 50) в своем геноме. PERVлатентно инфицирует многие культуры клеток свиньи и, реже, человека. Патогенность этого вируса не установлена.
Онковирусы, в отличие от лентивирусов, реплицируются в делящихся клетках. В составе ретровирусов обнаружено 11-13 белков. Часть из них локализована в сердцевине, а главный внутренний белок Р27 - Р30 - в мембране, окружающей РНК. В составе липопротеиновой оболочки 2-5 гликопротеинов, из которых более крупный образует головку шипика и является главным компонентом оболочки, а более мелкие белки образуют отросток, погруженный в билипидный слой. Различные белки ответственны за групповую и типовую специфичность. Подгрупповые и типоспецифические антигены связаны с наружными гликопротеинами и выявляются в РН, РСК, ИФ и РИД. Группоспецифические антигены связаны с белками сердцевины. Их обнаруживают в РСК, ИФ и РИД. Главный внутренний белок Р27 - РЗО - основной группоспецифический антиген, общий для всех ретровирусов птиц или млекопитающих, относящихся к одной группе.
Согласно предложенной ранее номенклатуре структурные белки ретровирусов обозначали следующим образом: МА - матриксный, СА - капсидный, NC -белок нуклеокапсида, SU - поверхностный, ТМ - трансмембранный, PR- протеаза, RT - ревертаза, IN - белок интеграции.