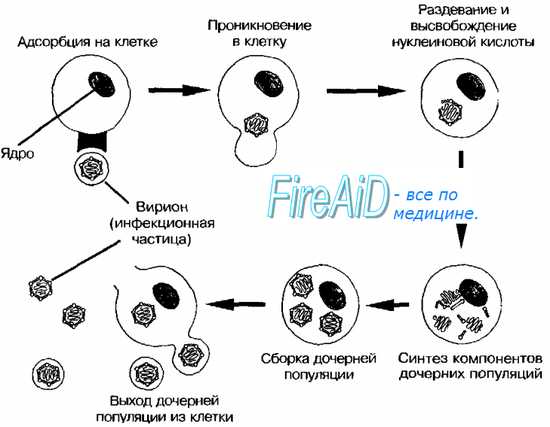
Заражение клеток вирусом. Зараженные вирусом клетки. Основные реакции зараженных вирусом клеток.
В заражённых вирусом клетках возможны патологические проявления разностороннего характера.
Морфология патологических эффектов. Зараженные вирусом клетки. Основные реакции зараженных вирусом клеток.
Характерные проявления взаимодействий между вирусом и чувствительными клетками-видимые поражения заражённых клеток вплоть до их гибели, а также присутствие возбудителей в исследуемом материале.
Альтерация и воспаление зараженных вирусом тканей. При вирусных инфекциях на первый план выступает картина повреждения клеток и воспалительных изменений тканей, при различных инфекциях их соотношение и выраженность варьируют. В противоположность бактериальным инфекциям (где доминируют полиморфноядерные лейкоциты), при вирусных поражениях среди клеточных элементов воспалительных реакций доминируют мононуклеары (лимфо- и моноциты). На этапах, предшествующих разрушению клеток, можно визуально наблюдать их дегенеративные и некротические изменения.
Форма зараженных вирусом клеток. Вследствие поражения цитоскелета клетки принимают округлую форму. Изменения структуры ядра достаточно разнообразны: кариопикноз (сморщивание ядра клетки при дистрофических изменениях в ней (от греч. karion, ядро, + pyknosis, уплотнение)], краевое расположение глыбок хроматина, его распыление и т.д. Подобные поражения способны вызывать адено-, герпес-, парамиксо-, ортомиксо- н ретровирусы. Плазмолемма (ЦПМ). Изменение ее структуры связано с замещением собственных гликопротеинов вирусиндуцированными белками. Эти поражения характерны для инфекций, вызванных оболочечными вирусами, но некоторые «голые» вирусы также могут вызвать изменения клеточной мембраны. Нарушение структуры плазмолеммы может приводить к изменениию некоторых её характеристик (например, к увеличению её проницаемости или слиянию с незаражёнными клетками). В последнем случае можно наблюдать образование симпластов (поли-кариоцитов) и синцитиев. Симпласты представлены гигантскими многоядерными клетками (например, клетки Цапка, выявляемые при герпетических поражениях), образующимися в результате модификации ЦПМ лизосомальными ферментами. Реже наблюдают образование синцитиев— больших конгломератов цитоплазмы, содержащих сотни и тысячи ядер связанных между собой клеток. Образование синцитиев обусловлено модификацией ЦПМ поверхностными гликопротеинами и характерно для парамиксовирусов.
Тельца включений зараженных вирусом клеток. Микроскопия заражённых клеток часто позволяет выявить тельца включений — характерный, но не абсолютный признак вирусных поражений. Тельца значительно крупнее, чем отдельные вирионы, и часто окрашиваются кислыми красителями (например, эозином).
• В одних случаях (например, при натуральной оспе) включения играют роль «вирусных фабрик», где собираются дочерние популяции, в других — служат депо побочных продуктов (например, при герпесвирусных инфекциях).
• При заражении клеток ДНК-содержащими вирусами тельца включений располагаются в ядре; исключение — тельца включений поксвирусов (тельца Гварнери).
• При заражении клеток РНК-содержашими вирусами тельца включений располагаются в цитоплазме (например, тельца Бабеша-Нёгри, выявляемые в цитоплазме клеток головного мозга при бешенстве).
Причины гибели зараженных вирусом клеток. Размножаясь в клетке, вирусы индуцируют синтез вирусспецифических белков, в той или иной степени подавляющих метаболизм клетки. Нарушение синтеза макромолекул вызвано нарушением трансляции клеточной мРНК. Среди РНК-геномных вирусов наиболее быстрое и глубокое подавление макромолекулярных синтезов в клетке вызывают пикорнавирусы, среди ДНК-геномных — покс- и герпесвирусы. Действие указанных вирусов реализуется на ранних этапах (до появления морфологических признаков цитопатического эффекта). Ингибирование синтеза РНК и ДНК обычно вторично по отношению к воздействию на белки, контролирующие экспрессию генов и пролиферацию клетки. Значительно реже нарушения вызывают вирусные белки, напрямую ингибирующие синтез нуклеиновых кислот. Среди РНК-геномных вирусов наиболее быстрое и глубокое подавление синтезов нуклеиновых кислот вызывают пикорнавирусы, среди ДНК-геномных — покс- и герпесвирусы.
Во время репродукции вируса в клетке накапливаются вирусные компоненты, оказывающие токсическое и повреждающее действие на клеточные структуры. Например, цитотоксические свойства проявляют капсомеры некоторых аденовирусов, гликопротеины парамиксовирусов. В процессе вирусной инфекции также происходит повреждение мембран лизосом, содержимое которых высвобождается и осуществляет аутолиз клетки. Таким образом, гибель клеток наступает в результате сочетания раннего подавления синтеза клеточных компонентов, накопления токсических вирусных продуктов и повреждения лизосом.