Было показано, что концентрация сахарозы в ситовидных трубках листьев обычно составляет от 10 до 30%, тогда как в фотосинтезирующих клетках она держится на уровне 0,5%. Следовательно, как мы уже отмечали, загрузка флоэмы идет против осмотического градиента. Этот механизм активно изучался в последние годы. Сначала растворенные органические вещества должны переместиться из хлоропластов во флоэму; этот путь составляет не более 3 мм, причем вещества могут двигаться и по симпласту, и по апопласту.
Симпластный путь предполагает перемещение веществ по плазмодесмам, а апопластный — через клеточные стенки посредством диффузии или объемного потока в транспирационном паре.
В 1968 г. Гуннинг и его сотрудники описали новый тип модифицированных клеток-спутниц — передаточные клетки. Как видно из рисунка, эти клетки примыкают к ситовидным трубкам. В результате дополнительного неравномерного утолщения клеточные стенки у них образуют многочисленные внутренние выступы, что почти в десять раз увеличивает площадь выстилающей такую стенку плазмалеммы. Как полагают, такое видоизменение клеток связано с необходимостью активно поглощать растворенные вещества из соседних клеток. Энергию для всех активных процессов дают многочисленные митохондрии передаточных клеток. Передаточные клетки обнаружены не у всех растений, но они обычны у бобовых и некоторых других семейств. Впрочем, считают, что активный транспорт происходит и в их отсутствие.
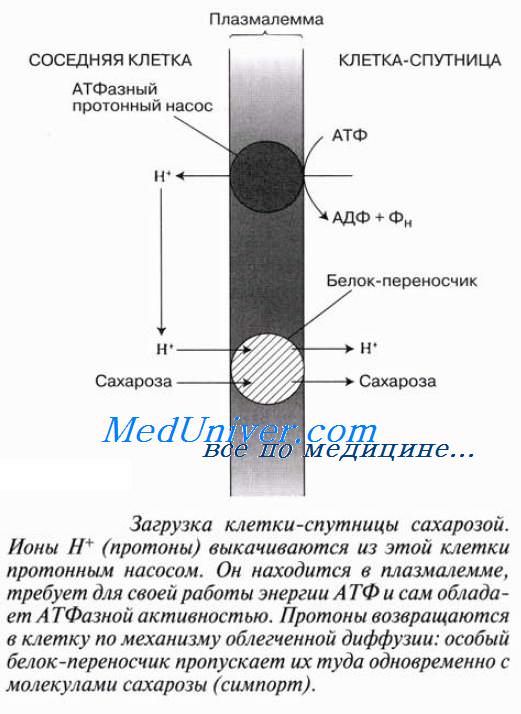
Как полагают, загрузку сахарозы (и ряда других компонентов раствора, в том числе аминокислот, фосфатов, ионов калия и аммония) в клетки-спутницы осуществляют особые белки-переносчики, локализованные в плазмалемме этих клеток. Образуемые белками системы переноса сходны с аналогичными системами животных и бактериальных клеток, где транспорт органических молекул сопряжен с транспортом протонов (Н+). Протоны выкачиваются из клетки переносчиком, использующим для этого энергию АТФ. В результате формируется градиент концентрации Н+, генерирующий потенциальную энергию. Протоны быстро диффундируют назад в клетку с участием других белков-переносчиков, которые одновременно пропускают внутрь молекулы сахарозы или других органических веществ (котранспорт по типу сим-порта). Таким образом, движущая сила этого процесса — формирование протонного градиента через плазмалемму с пониженным рН (более высокой концентрацией Н+) вне клетки.
В результате активного транспорта веществ в клетки-спутницы осмотический потенциал в них сильно понижается. Это стимулирует поступление в них воды путем осмоса, повышение давления и движение растворов (в том числе и раствора сахарозы) по механизму объемного потока через многочисленные плазмодесмы, связывающие клетки-спутницы с ситовидными трубками. Таким образом высокое давление и объемный поток создаются в ситовидных трубках, а не в клетках мезофилла листа, как предполагал Мюнх. Возможно также, что механизм активного транспорта через плазмодесмы из клеток-спутниц в ситовидные трубки точно такой же, как и механизм, обеспечивающий поступление сахарозы в клетки-спутницы.
При разгрузке ситовидных трубок растворенные вещества, вероятно, уходят из них в передаточные клетки по плазмодесмам. В результате водный потенциал трубок повышается (становится менее отрицательным). Это тормозит потенциальный осмос извне и поддерживает гидростатический градиент между местами загрузки и разгрузки флоэмы. Полагают, что последний процесс может идти и через плазмалем-му ситовидных трубок в их клеточные стенки и далее апопластным путем.