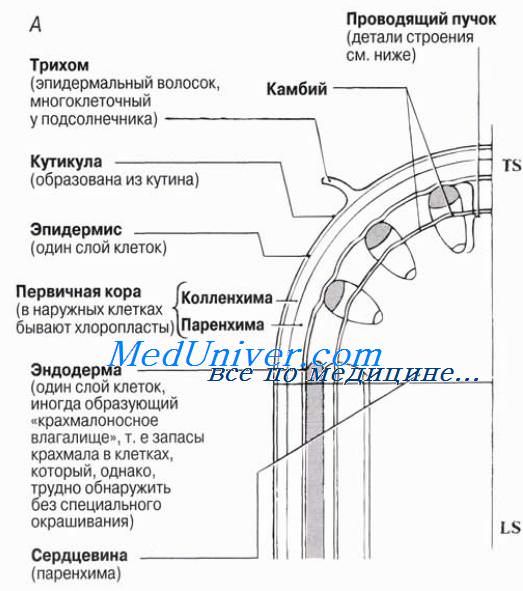
Ксилема цветковых растений содержит два типа проводящих воду структур — трахеиды и сосуды. В статье мы уже говорили о том, как выглядят эти структуры в световом микроскопе, а также на микрофотографиях, полученных с помощью сканирующего электронного микроскопа. Строение вторичной ксилемы (древесины) рассматривается в статье. Ксилема и флоэма образуют проводящую ткань высших, или сосудистых, растений. Эта ткань состоит из так называемых проводящих пучков, строение и распределение которых в стеблях двудольных растений с первичным строением показано на рисунке.
То, что вода поднимается именно по ксилеме, нетрудно продемонстрировать, погрузив побег срезанным концом в разбавленный водный раствор красителя, например эозина. Подкрашенная жидкость, распространившись вверх по стеблю, заполнит сеть пронизывающих листья жилок. Если затем сделать тонкие срезы и рассмотреть их в световом микроскопе, то окажется, что краситель находится в ксилеме.
Более эффектное доказательство подъема воды по ксилеме дают опыты с «кольцеванием». Такие опыты проводили задолго до того, как стали применяться радиоактивные изотопы, позволяющие очень легко проследить путь веществ в живом организме. В одном из вариантов опыта с одревесневшего стебля снимают узкое кольцо коры вместе с лубом, т. е. флоэмой. Довольно долго после этого находящиеся выше вырезанного кольца побеги продолжают расти нормально: следовательно, такое кольцевание не влияет на подъем воды по стеблю. Однако, если, приподняв лоскут коры, вырезать из-под него сегмент древесины, т. е. ксилемы, то растение быстро завянет. Таким образом, вода движется в побеги из почвы именно по этой проводящей ткани.
Любая теория, объясняющая транспорт воды по ксилеме, не может не учитывать следующие наблюдения.
1. Анатомические элементы ксилемы — тонкие мертвые трубки, диаметр которых варьирует от 0,01 мм в «летней» древесине до 0,2 мм в «весенней» древесине.
2. Большие количества воды движутся по ксилеме с относительно высокой скоростью: у высоких деревьев она составляет до 8 м/ч, а у других растений — около 1 м/ч.
3. Для подъема воды по таким трубкам к вершине высокого дерева необходимо давление порядка 4000 кПа. Самые высокие деревья — секвойи в Калифорнии и эвкалипты в Австралии — достигают в высоту более 100 м. Вода способна подниматься по тонким смачивающимся трубкам благодаря своему высокому поверхностному натяжению (это явление называется капиллярностью), однако только за счет этих сил даже по самым тончайшим сосудам ксилемы вода не поднимается выше 3 м.
Удовлетворительное объяснение этим фактам дает теория сцепления (когезии), или теория натяжения. Согласно этой теории, подъем воды от корней обусловлен ее испарением клетками листа. Как мы уже говорили в статье, испарение снижает водный потенциал клеток мезофилла, прилежащих к ксилеме, и вода поступает в эти клетки из ксилемного сока, водный потенциал которого выше; при этом она проходит через влажные клеточные стенки у концов жилок, как показано на рисунке.
Ксилемные сосуды заполняет сплошной столб воды; по мере того как вода выходит из сосудов, в этом столбе создается натяжение; оно передается вниз по стеблю до самого корня благодаря сцеплению (когезии) молекул воды. Эти молекулы стремятся «прилипнуть» друг к другу, потому что они полярны и притягиваются друг к другу электрическими силами, а затем удерживаются вместе водородными связями. Кроме того, они притягиваются к стенкам ксилемных сосудов, т. е. происходит их адгезия (прилипание) к ним. Сильная когезия молекул воды означает, что ее столб трудно разорвать — у него высокий предел прочности при растяжении. Растягивающее напряжение в клетках ксилемы приводит к генерированию силы, способной сдвигать весь водяной столб вверх по механизму объемного потока. Снизу вода поступает в ксилему из соседних клеток корня. При этом очень важно, что стенки ксилемных элементов жесткие и не спадаются при падении давления внутри, как это бывает, когда сосешь коктейль через мягкую соломинку. Жесткость стенок обеспечивается лигнином. Доказательством того, что жидкость внутри ксилемных сосудов сильно напряжена (растянута), служат суточные колебания диаметра древесных стволов, измеряемые инструментом под названием дендрограф.
Минимальный диаметр отмечен днем, когда интенсивность транспирации наивысшая. Натяжение столба воды в ксилемном сосуде немного втягивает внутрь его стенки (из-за адгезии), и сочетание этих микроскопических сжатий дает фиксируемую прибором общую «усадку» ствола.
Оценки прочности на разрыв столба ксилемного сока варьировали от 3000 доЗО 000 к Па, причем более низкие значения получены позднее. В листьях зарегистрирован водный потенциал порядка —4000 кПа, и прочность столба ксилемного сока, вероятно, достаточна, чтобы выдержать создающееся натяжение. Не исключено, конечно, что столб воды может иногда разрываться, особенно в сосудах большого диаметра.
Критики изложенной теории подчеркивают, что любое нарушение непрерывности столба сока должно немедленно останавливать весь поток, так как сосуд заполнится воздухом и паром (явление кавитации). Кавитацию может вызвать сильное сотрясение, изгибание ствола, а также дефицит воды. Хорошо известно, что на протяжении лета содержание воды в стволе дерева постепенно снижается, древесина заполняется воздухом. Этим пользуются лесозаготовители, потому что такие деревья легче сплавлять. Однако разрыв водного столба в части сосудов слабо влияет на общую скорость объемного потока. Возможно, дело в том, что вода перетекает в параллельно проходящие сосуды или же обходит воздушную пробку, продвигаясь по соседним паренхимным клеткам и по стенкам. Кроме того, согласно расчетам, для поддержания наблюдаемой скорости потока вполне достаточно, чтобы в каждый момент времени функционировала хотя бы небольшая доля ксилемных элементов. У некоторых деревьев и кустарников вода перемещается лишь по более молодой наружной древесине, называемой заболонью. У дуба и ясеня, например, проводящую функцию выполняют в основном сосуды текущего года, а остальная часть заболони играет роль водного резерва. Новые ксилемные сосуды образуются на протяжении всего вегетационного периода, но главным образом в его начале, когда скорость водного потока максимальна.
Вторая сила, обеспечивающая движение воды по ксилеме, — корневое давление. Его можно обнаружить и измерить в тот момент, когда срезают крону, а штамб с корнями некоторое время продолжает выделять сок из сосудов ксилемы. Этот процесс подавляется ингибиторами дыхания, например цианидом, и прекращается при недостатке кислорода и понижении температуры. Работа такого механизма, по-видимому, обусловлена активной секрецией солей и других водорастворимых веществ в ксилемный сок. В результате его водный потенциал падает, и вода поступает в ксилему из соседних клеток корня путем осмоса.
Этот механизм создает гидростатическое давление порядка 100-200 кПа (в исключительных случаях 800 кПа); одного его для подъема воды по ксилеме обычно недостаточно, однако у многих растений оно, несомненно, способствует поддержанию ксилемного тока. У медленно транспирирующих травянистых форм этого давления вполне хватает, чтобы вызвать у них мутацию. Так называется выделение воды на поверхности растения1 в виде капель жидкости, а не пара. Все условия, тормозящие транспира-цию, например слабая освещенность и высокая влажность, способствуют гуттации. Она обычна у многих видов дождевых тропических лесов и часто наблюдается на кончиках листьев у всходов трав.