Особый интерес представляет физиология сокращения матки. Установлено что финальным исходом начала маточного сокращения является возрастание концентрации ионов кальция (Са2+) в клетках миометрия в ответ на действие уте-ротонина. Определенную роль играют "щелевые контакты" (gap junction) между клетками миометрия для обеспечения ею сокращения и течения родового процесса. Эти "щелевые контакты" проявляются при доношенной беременности и их количество увеличивается в родах (Garfield R.E.G., 1988; Ulstrom U., 1992), а затем они исчезают в течение 24 часов после родов. Эти "щелевые контакты" отсутствуют в миометрии при недоношенной беременности и наблюдаются при преждевременных родах и при родах спонтанных или индуцированных. В исследованиях на животных in vivo и in vitro установлено, что прогестерон предотвращает, а эстрогены способствуют образованию связей ("щелевых контактов")- Для этого необходим синтез протеина, важна также роль простагландинов, тромбоксана и, возможно, эндоперекиси. Угнетение синтеза ПГ подавляет образование "щелевых контактов".
Сокращения волокон миометрия отличаются от такового скелетной мускулатуры. Расположение волокон в миометрии в различном направлении способствует усилению выталкивающей силы матки независимо от положения и предлежания плода.
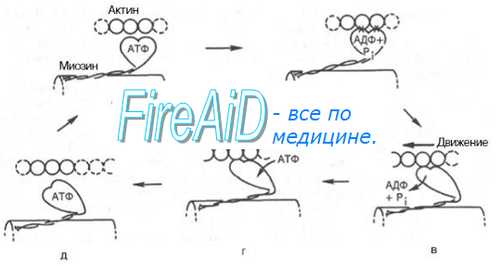
Молекулярная регуляция гладкомышечного сокращения. В мышечном сокращении основную роль играет миозин. У молекулы миозина есть функциональные "головная" и "хвостовая" части. Шаровидная (глобулярная) "головная" часть является местом взаимодействия с актином, т.е. местом, где генерируется сила сокращения; местом, где находится АТФ-аза, под действием которой гидролизуется АТФ, и, тем самым, химическая энергия превращается в физическую силу; местом, где расположены легкие цепи миозина с низкой молекулярной массой («20 000), фосфорилирование которых представляет собой ключевую реакцию в регуляции сокращения, т.е. во взаимодействии актина и миозина в гладкой мышце.
В связи с этим важно помнить, что для генерации мышечного сокращения необходимо поступление ионов кальция (Са+) из внеклеточных пли внутриклеточных запасов. Са накапливается внутри клеток в пузырьках саркоплазматичес-кого ретикулума. В гладких мышцах взаимодействие актина и миозина peгулируется ферментативным фосфорилированием (или дефосфорилированием) легких цепей миозина (Still 1 J.T, et al., 1980). В клетках миометрия взаимодействие актина с миозином происходит только после фосфорилирования легкой цепи миозина. Оно катализируется ферментом киназа легкой цепи миозина; важно, что этот фермент активизируется Са2+. Соединения, которые увеличивают содержание внутриклеточного свободного Са, усиливают сокращения гладкой мускулатуры миометрия. Кальций связывается с кальмодулином (регуляторным белком, который опосредует эффект кальция), который в свою очередь, связывается с киназой легкой цепи миозина и активирует ее.
Ситуации, уменьшающие уровень свободного внутриклеточного кальция, способствуют расслаблению. Дефосфорилирование легкой цепи миозина под действием фосфатаз также способствует расслаблению мышцы. Актин не взаимодействует с нефосфорилированным миозином. Соединения, которые повышают внутриклеточную концентрацию циклического АМФ (цАМФ), усиливают релаксацию матки. Эти соединения включают р-адренергические агонисты. Считается, что цАМФ вызывает снижение внутриклеточного Са2, хотя это не доказано. Согласно другой точке зрения, происходит инактивация фермента киназа легкой цепи миозина в результате его фосфорилирования под воздействием цАМФ-зависи-мого фермента протеинкиназы.
Снижение активности фосфорилирования фермента объясняется снижением его сродства к кальмодулину; для проявления активности этого фермента его взаимодействие с кальмодулином должно быть обязательным.
Можно считать, что регуляция сократительной активности миометрия на клеточном уровне осуществляется при посредстве киназы легкой цепи миозина, па активность которой влияет кальцин. Одновременно следует учитывать и дефосфорилирование её под действием соответствующей фосфатазы. Активация сокращения осуществляется путем взаимодействия фосфорилированного миозина и актина с образованием фосфорилированного актомнозина.
Акт родов протекает при наличии сформированной родовой доминанты, объединяющей в динамическую систему как высшие нервные центры, так и исполнительные органы. В формировании родовой доминанты большое значение имеет воздействие половых гормонов па различные образования центральной и периферической нервной системы. Центральная нервная система осуществляет высшую и тонкую регуляцию родового акта. Значительное возрастание электрической активности головного мозга наступает за 1-1,5 нед до наступления родов.
Большое значение для возникновения родовой деятельности и правильного ее течения имеет подготовленность женского организма, готовность матки, а также чувствительность миометрия к воздействию контрактильных веществ.