Гормоны щитовидной железы ( T3, T4, кальцитонин) : их образование, транспорт, метаболизм и регуляция через ТТГ. Йод в организме человека
Нормальная функция щитовидной железы направлена на секрецию L-тироксина (Т4) и 3,5,3'-трийод-L-тиронина (Т3) - йодированных аминокислот, которые представляют собой активные тиреоидные гормоны и влияют на разнообразные метаболические процессы (рис. 324-1). Заболевания щитовидной железы проявляются качественными или количественными изменениями секреции гормонов, увеличением размеров органа (зоб) или тем и другим вместе. Недостаточность секреции гормонов приводит к развитию синдрома гипотиреоза, или микседемы, главной особенностью которого служит снижение калорических затрат (гипометаболизм). Напротив, чрезмерная секреция активных гормонов вызывает появление гиперметаболизма и других признаков синдрома, называемого гипертиреозом, или тиреотоксикозом. Увеличение массы щитовидной железы (составляющей у взрослого человека в норме 15-25 г) может быть диффузным или очаговым. Диффузное увеличение необязательно должно быть полностью симметричным. Обычно правая доля железы увеличивается больше, чем левая. Такое увеличение может сопровождаться повышенной, нормальной или сниженной секрецией гормонов, что определяется причиной заболевания. Истинно очаговое увеличение отражает, как правило, наличие новообразований, будь то добро- или злокачественных. Первые иногда обусловливают гиперсекрецию тиреоидных гормонов и гипертиреоз, тогда как при злокачественных опухолях это наблюдается очень редко. Зоб любого типа может привести к сдавлению соседних структур шеи и средостения.
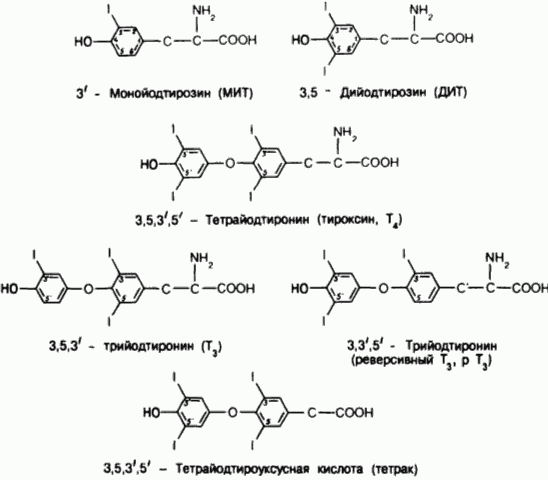
Структурные формулы тироксина, его предшественников и некоторых метаболитов.
Щитовидная железа человека развивается в эмбриогенезе из выпячивания глоточного эпителия и клеток латеральных глоточных карманов. Постепенно опускаясь по средней линии, зародышевая щитовидная железа формирует щитоязычный проток, который тянется от слепого отверстия у основания языка до перешейка железы. По ходу этого тракта могут сохраняться остатки ткани в виде "язычной щитовидной железы", щитоязычных кист и узлов или структуры, прилегающей к перешейку щитовидной железы и называемой пирамидальной долей. Последняя обычно видна только в случае увеличения остатка железы. У некоторых людей единственной функционирующей тиреоидной тканью может быть "язычная щитовидная железа", секреция которой бывает либо достаточной, либо недостаточной для сохранения нормального метаболического (эутиреоидного) статуса.
Аплазия щитовидной железы и функциональная недостаточность эктопической тиреоидной ткани служат причиной спорадического неонатального гипотиреоза, имеющего важное значение для здравоохранения вследствие частоты встречаемости (1 случай на 4000-5000 новорожденных) и способности отвечать на своевременно начатое лечение.
Щитовидная железа плода приобретает способность концентрировать и органифицировать йод примерно к 10-й неделе беременности. Вскоре после этого в крови уже удается определять как Т4, так и тиреотропный гормон (тиреотропин, ТТГ), концентрации которых на протяжении II триместра беременности непрерывно возрастают. Увеличение уровня Т4 в сыворотке крови плода обусловлено как усилением секреторной активности щитовидной железы и появлением в плазме тироксинсвязывающего глобулина (ТСГ), так и повышением уровня ТТГ, отражающим созревание гипоталамуса плода и секрекцию им тиреотропинрилизинггормона (ТРГ). Материнский ТРГ легко проникает через плаценту и, по-видимому, играет роль в развитии гипофизарно-тиреоидной системы плода.
Напротив, материнский ТТГ через плаценту не проникает. Т3 появляется в крови плода позднее, но также в течение II триместра, и его концентрация в крови и амниотической жидкости остается низкой вплоть до начала послеродового периода. В отличие от этого концентрация его аналога (реверсивного Т3, рТ3) в крови плода и амниотической жидкости превышает таковую в крови матери. Эти различия отражают качественные особенности метаболизма 'T4 у плода, которые рассматриваются ниже. Низкая концентрация Т3 в крови плода и амниотическое жидкости на фоне его высокой концентрации у матери свидетельствует о минимальном переносе Т3 от матери к плоду, что характерно и для Т4. Следовательно, основным тиреоидным гормоном, получаемым плодом, является Т4, продуцируемый собственной щитовидной железой плода. Таким образом, за исключением возможного влияния материнского ТРГ, гипофизарно-тиреоидная ось плода представляет собой функциональную систему, независимую от таковой у матери.
Нормальная щитовидная железа у взрослого человека состоит из соединенных перешейком двух долей и располагается кпереди и книзу от хрящей гортани. Фиброзные перегородки делят железу на псевдодольки, которые в свою очередь состоят из везикул, называемых фолликулами, или ацинусами, окруженных сетью капилляров. В норме стенки фолликула выстланы эпителиальными фолликулярными клетками кубической формы. Просвет фолликула заполнен белковым материалом, получившим название коллоид, который содержит специфический для щитовидной железы белок - тиреоглобулин, ответственный за синтез и накопление Т4 и Т3. В щитовидной железе присутствует и другая популяция клеток - С-клетки. Они служат источником кальцитонина, а их злокачественное перерождение приводит к медуллярному раку щитовидной железы.
Понятие "динамика тиреоидных гормонов" означает комплекс процессов синтеза гормонов в щитовидной железе, их транспорта в крови, действия и метаболизма в периферических тканях, а также комплекс регуляторных механизмов, определяющих нормальное обеспечение тканей тиреоидными гормонами.
Синтез тиреоидных гормонов зависит от поступления в щитовидную железу достаточного количества йода - составной части активных гормонов (Т4 и Т3), интактности путей метаболизма йода в железе и одновременного синтеза
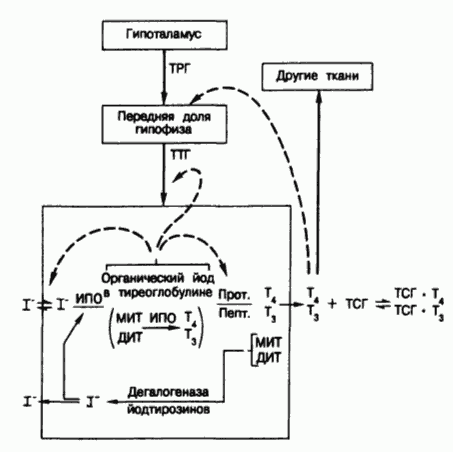
Схема путей синтеза и секреции тиреоидных гормонов и механизмов супра- и интратиреоидной регуляции функции щитовидной железы.
Тонкими стрелками показаны пути метаболизма йода; жирными стрелками - стимулирующие влияния; пунктиром - ингибирующие влияния.
Обозначения : ТРГ - тиреотропин-рилизинг гормон, ТТГ - тиреотропный гормон, И ПО - йодид-пероксидаза, Прот - тиреоидная протеаза, Пепт - тиреоидная пептидаза, МИТ - монойодтирозин, ДИТ - динодтирозин, T4 - тироксин, Т3 - 3, 5, 3' - трийодтиронин.
Секреция достаточного количества гормонов требует в свою очередь, как нормальной скорости их синтеза, так и интеграции с протекающими в железе процессами гидролиза тиреогло-булина, в результате которых активные гормоны высвобождаются. Йод проникает в щитовидную железу из крови в форме неорганического или органического йодида. Существует два источника его поступления: первый - при дейодировании тиреоидных гормонов или насыщенных йодом агентов, попавших в организм человека; и второй - с пищей, водой или лекарственными препаратами. Раньше для населения континентальной части США считалось нормой потребление с пищей примерно 200 мкг йода; этого было достаточно для поддержания концентрации йодида в плазме па уровне приблизительно 0,5 мкг/дл (5 мкг/л).
Однако из-за присутствия йода в некоторых пищевых продуктах и широкого распространения йодсодержащих лекарственных средств, витаминных препаратов и антисептиков среднее потребление йода возросло до 1000 мкг в сутки, что привело к соответствующему повышению концентрации йодида в плазме крови.
Йодид извлекается из плазмы щитовидной железой, почками, а также слюнными железами и в желудочно-кишечном тракте, но, поскольку йодид, вы-деляющийся в просвет кишечника, подвергается реабсорбции, чистый его клиренс осуществляется только щитовидной железой и почками. В сущности, щитовидная железа и почки конкурируют друг с другом за йодид плазмы. Почечный клиренс зависит в основном от скорости клубочковой фильтрации, и на него не влияют гуморальные факторы или концентрация йодида в плазме. Поэтому почки в норме являются пассивными участниками этой конкуренции.
Отсюда следует, что соотношение между скоростью поступления йодида в щитовидную железу и скоростью его экскреции с мочой определяется активностью именно щитовидной железы, а не почек.
Процессы синтеза и секреции активных тиреоидных гормонов можно разделить на четыре последовательных этапа. Первый включает активный транспорт йодида из плазмы в клетку щитовидной железы и в просвет фолликула. Скорость этого процесса превышает скорость пассивной днффузии йода из железы. В результате щитовидная железа оказывается способной удерживать градиент концентрации для йодида (отношение концентраций щитовидная железа/плазма) на весьма высоком уровне (до 500 и более в определенных условиях). Энергия для транспорта йодида черпается из фосфатных связей и поэтому зависит от окислительного фосфорилирования в железе. Второй этап биосинтеза гормонов включает окисление йодида в более ре-акционноспособную форму, способную йодировать тирозиновые остатки в молекуле тиреоглобулина - гликопротеида с мол. массой около 650 000, который синтезируется клетками фолликулов.
Окисление йодида осуществляется йодид-пероксидазой, использующей перекись водорода, которая образуется но ходу окислительного обмена в железе. Йодирование органических структур происходит на границе между клеткой и коллоидом, где этому процессу подвергается в основном свежесинтезированный тиреоглобулин, поступающий путем экзоцитоза в просвет фолликула. В результате в составе пептида образуются неактивные предшественники гормонов - монойодтирозин (МИТ) и дийодтирозин (ДИТ). Затем эти йодтирозины вступают в реакцию окислительной конденсации опять-таки с помощью пероксидазы. Данная реакция протекает внутри молекулы тиреоглобулина и приводит к образованию различных йодтиронинов, включая Т4 и Т3 Хотя в крови и присутствуют небольшие количества тиреоглобулина, большая его часть некоторое время хранится в железе, играя роль запасной формы тиреоидных гормонов, или "прогормона". Высвобождение активных гормонов в кровь происходит путем пиноцитоза фолликулярного коллоида на апикальном краю клетки с образованием коллоидных капелек. Для этого процесса необходимо функционирование микротрубочек.
Коллоидные капельки сливаются с тиреоидными лизосомами, образуя "фаголизосомы", в которых тиреоглобулин гидролизуется протеазами и пептидазами. Конечный этап заключается в выделении свободных йодтиронинов - Т4 и Т3 - в кровь. Единственным источником эндогенного Т4 служит щитовидная железа. В отличие от этого только около 20% образующегося в норме Т3 поступает из щитовидной железы; остальная его часть образуется во внетиреоидных тканях путем ферментативного отщепления 5'-йода от наружного кольца молекулы Т . Неактивные йодтиро-зины, высвобождающиеся при гидролизе тиреоглобулина, отдают свой йод под действием внутритиреоидного фермента - дегалогеназы йодтирозинов. В норме высвобождающийся таким образом йод в основном реутилизируется в синтезе гормонов, но небольшая его доля все же теряется, поступая в кровоток ("утечка йода"). В патоло-гических условиях эта доля может возрастать.
Щитовидная железа способна концентрировать и другие одновалентные анионы, такие как пертехнетат, который имеется в виде радиоактивного изотопа - натрий [99mТc] пертехнетат. В отличие от йодида пертехнетат очень мало связывается орга-ническими соединениями. Поэтому он присутствует в щитовидной железе только короткое время. Это свойство наряду с его коротким физическим периодом полураспада делает пертехнетат ценным радионуклидом для получения изображения щитовидной железы с помощью методов сцинтилляционного сканирования.
Перечисленные выше реакции служат объектом торможения различными химическими соединениями. Их обычно называют зобогенными веществами, поскольку в силу своей способности ингибировать синтез гормонов и косвенно стимулировать секрецию ТТГ они вызывают образование зоба. Некоторые неорганические анионы, в том числе перхлорат и тиоцианат, ингибируют механизм транспорта йодида и тем самым уменьшают доступность субстрата для образования гормонов. Однако развивающиеся в результате этого зоб и гипотиреоз можно предотвратить или ликвидировать достаточно большими дозами йодида, которые обеспечивают поступление нужных его количеств в железу за счет простой диффузии. Широко используемые антитиреоидные средства, такие как производные тиомочевины и меркаптоимидазола, оказывают на биосинтез гормонов более сложное воздействие.
Эти вещества, равно как и некоторые производные анилина, ингибируют первоначальное окисление (органическое связывание) йодида, снижая долю образующегося ДИТ относительно МИТ и блокируя конденсацию йодтирозинов в гормонально-активные йодгиронины. Последняя реакция наиболее чувствительна. Таким образом, синтез гормонально-активных йодтиронинов может быть резко заторможен в условиях лишь небольшого снижения общего захвата йода щито-видной железой. В отличие от эффекта одновалентных анионов зобогенное действие ингибиторов органического связывания йода не преодолевается большими его количествами. Действительно, некоторые слабые зобогенные вещества, такие как сульфона-миды и антипирин, при введении вместе с йодидом становятся почему-то даже более активными. Острое введение больших доз самого йода тоже может приводить к блокаде органического связывания и реакции конденсации. В норме это действие (эффект Вольффа - Чайкоффа) транзиторно, но у некоторых здоровых лиц, длительно получаю-щих йод, имеет место постоянное торможение синтеза гормонов, сопровождающееся развитием зоба с гипотиреозом (йодная микседема) или без него.
Большинство больных с болезнью Грейвса, особенно перенесшие радиойодтерапию или хирургическую операцию, а также больные с болезнью Хашимото чрезвычайно чувствительны к блокирующему действию йодида, и при хроническом приеме йодидов у них развивается гипотиреоз. Точно так же высокую чувствительность обнаруживает и щитовидная железа плода, и поэтому во избежание зобного гипотиреоза у плода беременные женщины не должны получать больших доз йодида. Йодид в больших дозах может ингибировать и протеолиз тиреоглобулина, т. е. высвобождение гормонов. Этот эффект легче всего проявляется в условиях гиперфункции щитовидной железы, и именно он определяет быстрое терапевтическое действие йодидов у большинства больных гипертиреозом. Литий, вводимый ряду больных с депрессивными состояниями в виде карбонатной со-ли. оказывает несколько эффектов на внутритиреоидный обмен йода, один из которых заключается в торможении секреции гормонов. Большие дозы дексаметазона также ин-гибируют секрецию гормонов и в сочетании с йодидом могут быстро уменьшать выра-женность тиреотоксикоза.
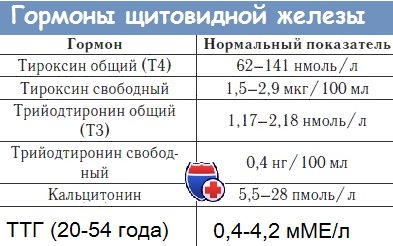
Гормоны щитовидной железы ( T3, T4, кальцитонин): Транспорт и метаболизм гормонов
Транспорт гормонов. В крови Т4 и Т3 почти полностью связаны с белками плазмы. В порядке уменьшения интенсивности связывания Т4 эти белки располагаются следующим образом: альфа-глобулин, называемый тироксин- или тиронин-связывающмм глобулином (ТСГ), Т4-связывающий преальбумин (ТСПА) и альбумин. Из-за своего вы-сокого сродства к Т4 СТГ в норме является главной детерминантой общей связывающей активности плазмы. Взаимодействие между Т4 и его связывающими белками формирует обратимое равновесное состояние, при котором большая часть гормона оказывается связанной, а очень малая его доля (в норме около 0,03%) - свободной. Т3 связывается с ТСПА в незначительной степени, а с ТСГ- менее прочно, чем Т4. Вследствие этого доля свободного Т3 в норме (примерно 0,3%) в 8-10 раз превышает таковую свободного Т4.
Ткани используют только свободный, или несвязанный, гормон. Поэтому метаболический статус теснее коррелирует с концентрацией именно свободного гормона, чем с общей его концентрацией в плазме, а гомеостатическая регуляция тиреоидной функции тоже направлена на поддержание нормальной концен-трации свободного, а не общего гормона. Кроме того, относительно слабое связывание Т3 обусловливает незначительность его вклада в общую концентрацию белково-связанного гормонального йода в крови и, возможно, более быстрое начало и окончание его действия. Нарушения взаимодействия между тиреоидными гормонами и бел-ками плазмы бывают двух общих типов.
Таблица. Классификация различных нарушений взаимодействия тиреоидных гормонов с белками плазмы
Тип нарушения
Уровень Т4 и Т3 в сыворотке крови
Процент СТ4 и СТ3 или ПТ3С
СТ4 и СТ3 или ИСТ4 и СТ3
I. Первичное изменение ТСГ
Повышенная концентрация
повышается
снижается
Н
Сниженная концентрация
снижается
повышается
Н
II. Первичное нарушение функции щитовидной железы
Гипотиреоз
снижается
Гипертиреоз
повышается
повышается
повышается
Обозначения: СТ4 - свободный Т4; СТ3- свободный Т3; ИСТ4 - индекс свободного Т4; ИСТ3-индекс свободного Т3; ПТ3С-поглощение Т3 смолой; ТСГ-тироксинсвязывающий глобулин.
Повышенный уровень ТСГ
Сниженный уровень ТСГ
Беременность
Неонатальный период
Пероральные контрацептивы и другие источники эстрогенов
Тамоксифен
Инфекционный и хронический активный гепатит
Билиарный цирроз Острая интермиттирующая порфирия
Перфеназин
Генетические причины
Андрогенные и анаболические стероиды Большие дозы глюкокортикоидов
Хронические заболевания печени Активная акромегалия
Некроз
Генетические причины
Аспарагиназа
Андрогенные и анаболические стероиды
Большие дозы глюкокортикоидов
Хронические заболевания печени
Активная акромегалия
Некроз
Генетические причины
Аспарагиназа
В первом случае ось щитовидная железа - гипофиз не нарушена, и гомеостатическая регуляция секреции тиреоидных гормонов сохраняется. В таких условиях нарушение взаимодействия обусловливается изменением связывания тиреоидных гормонов. Например, увеличение уровня ТСГ вначале снижает концентрацию свободного гормона и тем самым уменьшает его доступность для тканей. Затем общая концентрация гормона в сыворотке возрастает до тех пор, пока концентрация свободного гормона не восстановится до нормы. При этом доли свободных Т4 и Т3 снижаются. Увеличение общей концентрации гормона уравновешивает снижение доли свободной его формы и в результате абсолютная концентрация свободного гормона остается нормальной, что определяет и нормальный метаболический статус.
При снижении концентрации ТСГ происходят противоположные изменения. Состояния, сопровождающиеся первичными сдвигами в концентрации ТСГ, перечислены в таблице. Первичные нарушения связывания тиреотропных гормонов происходят при увеличении содержания в плазме и других связывающих белков, а также при появлении патологических связывающих белков. Эти вопросы обсуждаются ниже.
Во втором случае нарушение связывания тиреоидных гормонов обусловлено первичными изменениями их концентрации в крови, как это характерно для гипотиреоза или тиреотоксикоза. При этом нормальная гомеостатическая регуляция секреции тиреоидных гормонов теряется либо из-за нарушения самих регуляторных механизмов, либо потому, что интактные регуляторные механизмы оказываются неспособными преодолеть эффекты какой-либо патологии вне гомеостатической системы. В таких условиях концентрация ТСГ почти не меняется, а концентрация свободного гормона ока-зывается прямо пропорциональной его общей концентрации. Поскольку гомеостатиче-ские механизмы не могут восстановить нормальную концентрацию свободного гормона, первичные нарушения функции щитовидной железы сопровождаются постоянными измене-ниями концентрации общего и свободного гормона и, следовательно, изменениями ме-таболического статуса. При таких нарушениях доля свободного гормона меняется в том же направлении, что и поступление гормона в кровь.
После своего проникновения в клетку Т4 и Т3 вступают в различные реакции, которые в конце концов приводят к их экскреции или инактивации. Метаболизм тиреоидных гормонов сводится главным образом к последовательному удалению каждого атома йода (монодейодирование) с образованием в конечном счете полностью лишенного йода тиронинового ядра.
Дейодированню подвергаются примерно 70% Т4 и Т3. В случае Т4 наибольшую важность имеет 5'-монодейодирование, которое приводит к образованию Т3 (Т3-неогенез). Поскольку около 30% Т4 превращается в Т3 и поскольку Т3 обладает примерно втрое большей метаболической активностью, чем Т4, практически весь метаболический эффект Т4 может быть отнесен на счет образующегося из него Т3. В нормальных условиях Т3-неогенез определяет примерно 80% присутствующего в крови Т3 и его общей продук-ции; остальное количество непосредственно секретируется щитовидной железой. По-этому патологические состояния и фармакологические средства, которые нарушают Т3-неогенез, понижают концентрацию Т3 в сыворотке.
Когда больные с гипофункцией щитовидной железы получают такие дозы синтетического Т4 (левотироксина), которые поддерживают его концентрацию в сыворотке на нормальном или слегка повышенном уровне, в крови создается нормальная или почти нормальная концентрация Т3.
Состояния, сопровождающиеся снижением периферической конверсии Т4 в Т3:
I. Физиологические:
Эмбриональный и ранний неонатальный период
Старческий возраст
II. Патологические:
Голодание
Нарушение питания
Системные заболевания
Физическая травма
Послеоперационный период
Фармакологические средства (пропилтиоурацил, дексаметазон, пропранолол, амиодарон)
Рентгеноконтрастные средства (ораграфин, телепак)
Положение о том, что щитовидная железа секретирует сравнительно небольшие количества Т3, неприменимо к состояниям, при которых имеет место автономная гиперфункция щитовидной железы, избыточная ее стимуляция ТТГ или сниженное содержание йода в ней. В таких условиях отношение Т3/Т4 в продуктах секреции щитовидной железы и в крови увеличивается.
Кроме того, при сниженной продукции Т4, как это наблюдается на ранних стадиях тиреоидной недостаточности или при дефиците йода, отношение концентраций Т3/Т4 в крови возрастает еще больше в ре-зультате срабатывания ауторегуляторного механизма, повышающего эффективность Т3-неогенеза.
Примерно 40% Т4 подвергается монодейодированию в положении 5 внутреннего кольца с образованием 3,3', 5'-трийод-L-тиронина (реверсивный Т3, рТ3). Этот процесс определяет почти всю продукцию рТ3 в организме. Реверсивный Т3 практиче-ски не обладает метаболической активностью. Поэтому соотношение между процессами монодейодирования наружного и внутреннего колец детерминирует количество доступ-ного для тканей метаболически активного гормона. Факторы, нарушающие Т3-неогенез, почти всегда увеличивают концентрацию рТ3 в сыворотке.
Это увеличение связано не с повышенной продукцией рТ3 из Т4, а с торможением 5'-монодейодирования рТ3, в результате которого образуется 3,3'-дийодтиронин (3,3'-Т3). Иными словами, как снижение конверсии Т4 в Т3, так и снижение деградации рТ3 обусловлено избирательным нарушением 5'-монодейодирования.
Второй главный путь метаболизма Т4, Т3 и их метаболитов заключается в их конъюгировании в основном с глюкуронатом и сульфатом в печени. Эти конъюгаты либо подвергаются дейодированию на месте, либо выделяются в желчь, но размеры энтерогепатического кругооборота у человека неизвестны. Даже в лучшем случае про-исходит неполная реабсорбция и на долю экскреции Т4, Т3 и их йодсодержащих метаболитов с калом приходится примерно 20% общей элиминации Т4. Небольшая доля Т4 и Т3 (около 20%) подвергается окислительному дезаминированию и декарбоксилированию по боковой цепи аланина с образованием уксуснокислых аналогов - тетрайод- и трийодтироуксусных кислот (тетрак и триак соответственно).
В некоторых условиях изменения скоростей метаболического клиренса Т4 и Т3 определяются в основном сдвигами в накоплении и метаболизме гормонов. Фенобарбитал и фенитоин ускоряют метаболический клиренс тиреоидных гормонов, не увеличивая долю свободных гормонов в крови. Больше того, что касается фенитоина, то он снижает концентрацию общего и свободного Т4. Тем не менее нормальный метаболический статус поддерживается, вероятно, за счет повышения Т3-неогенеза.
Гормоны щитовидной железы ( T3, T4, кальцитонин): Действие гормонов.
Тиреоидные гормоны влияют на рост и созревание тканей, общие энергозатраты и кругооборот практически всех субстратов, витаминов и гор-монов, включая и сами тиреоидные гормоны. Первичные механизмы возникновения этих эффектов остаются неясными, но, по-видимому, гормоны действуют координирование на уровне клеточного ядра (изменяя экспрессию генома), на уровне митохондрий (влияя на окислительный обмен) и на уровне плазматической мембраны (регулируя поток субстратов и катионов в клетку и из нее).
Видео гормоны щитовидной железы в норме и при болезни